


BIO – Medicina Costruzione Sociale nella Post-Modernità – Educational Papers • Anno V • Numero 18 • Giugno 2016
Scritto in collaborazione con Eugenia D’Alterio – biologa
I mitocondri nella cultura popolare
Descrivere i mitocondri sarebbe una questione facile se ci limitassimo a dire che sono le centrali energetiche delle cellule eucariote sia animali che vegetali. I mitocondri come i cloroplasti (presenti solo nelle cellule vegetali e preposti alla fotosintesi clorofilliana) sono organuli citoplasmatici che secondo la teoria endosimbiotica si sono evoluti da batteri vissuti all’interno di cellule più specializzate, ossia le prime cellule eucariote. Arrivare a tale descrizione e classificazione è stato, tuttavia, un percorso di costruzione sociale di conoscenza controverso. La complessità che racchiude questa semplice descrizione dei mitocondri (così come per i cloroplasti) è anche essa controversa, innanzitutto per le intuizioni circa la natura e l’evoluzione della vita stessa che la questione dei mitocondri continua a porre agli studiosi.
Cosi, dopo un secolo di ricerca sui mitocondri1, si può postulare che essi sono minuscoli organelli all’interno delle cellule che producono quasi tutta la nostra energia sotto forma di ATP2 . In media ci sono migliaia di mitocondri in ogni cellula umana, per cui si stima che nel corpo umano ce ne siano circa dieci milioni di miliardi. In sostanza, tutte le cellule complesse (animali e vegetali) contengono mitocondri. Questi organuli citoplasmatici una volta erano batteri che si adattarono alla vita all’interno delle cellule più specializzate (le prime cellule eucariote), circa due miliardi di anni fa. Essi conservano un frammento di genoma come vestigia della loro precedente indipendenza. I loro rapporti con le loro cellule ospiti hanno plasmato l’intero tessuto della vita, dall’energia al sesso, alla fertilità, al suicidio delle cellule2 e al loro invecchiamento e alla loro morte3.
Molte persone hanno sentito parlare dei mitocondri per un motivo o per un altro, anche dal punto di vista forense. In alcuni giornali e libri di testo, sono sommariamente descritti come le “centrali energetiche della vita”4, cioè come piccoli generatori all’interno delle cellule che producono praticamente tutta l’energia di cui abbiamo bisogno per vivere (i processi mitocondriali garantiscono la respirazione cellulare adempiendo alla scissione del glucosio in presenza di ossigeno in CO2 e H2O. Ce ne sono, solitamente, migliaia in una singola cellula, in cui questi organuli usano ossigeno per bruciare il glucosio. Sono così piccoli che un miliardo di loro potrebbe stare facilmente in un granello di sabbia5.
Da ricordare che ogni mitocondrio ha del DNA (denominato anche come mtDNA) che viene ereditato esclusivamente dalla madre, la linea materna è fondamentale nell’eredità di questo DNA (da non confondersi con il DNA cromosomico nucleare, il DNA più comunemente usato per eseguire i test come il test di paternità, o test per identificare una persona, in ambito forense, per via del fatto che risulta molto più resistente all’impatto di fattori ambientali, come contaminazioni o quando si tratta di identificare un corpo bruciato, l’analisi del mtDNA si presenta come un alternativa valida all’analisi del DNA nucleare tradizionale. Infatti il DNA nucleare si degrada abbastanza facilmente se non conservato correttamente).
Alcune persone hanno familiarità con l’espressione “Eva mitocondriale”, presumibilmente l’antenato più recente comune a tutti i popoli d’oggi, se rintracciassimo il nostro patrimonio genetico, risalendo la linea materna, dalla madre alla nonna materna e così via, risalendo nelle profonde nebbie del tempo. Questa teoria propone che l’Eva mitocondriale, la madre di tutte le madri, conosciuta anche come l’Eva africana, abbia vissuto in Africa, forse 170.000 anni fa. Secondo questa teoria, con questa modalità, noi possiamo rintracciare la nostra origine genetica poiché tutti i mitocondri citoplasmatici hanno mantenuto una piccola quota dei loro geni, che di solito sono trasmessi alla generazione successiva solo nella cellula uovo, ma non nei gameti maschili. Ciò significa che i geni mitocondriali si comportano come un nome del genere femminile, che ci permette di rintracciare i nostri antenati lungo la linea materna nello stesso modo in cui alcune famiglie cercano di tracciare la loro ascendenza lungo la linea maschile (genoma nucleare tramite il cromosoma Y) fino a Arduino Visconte di Narbona, a Noah, o a Maometto6. Recentemente, alcuni di questi postulati sono stati messi in discussione ma, in generale, la teoria rimane una valida risorsa euristica. Naturalmente, la teoria non solo dà un’idea dei nostri antenati ma, anche, aiuta a chiarire chi non lo era. Secondo l’analisi dei geni mitocondriali materni, l’uomo di Neanderthal non si incrociò con il moderno Homo sapiens, ma è stato portato all’estinzione ai margini del territorio che oggi consideriamo Europa7.
I mitocondri hanno anche fatto i titoli dei giornali per il loro uso in medicina legale, essendo più stabile rispetto al DNA nucleare come già accennato, per stabilire l’identità di persone o cadaveri, in diversi casi celebri. In questo caso, la tecnica si basa sulla piccola quota di geni mitocondriali ma perfettamente integri. L’identità dell’ultimo zar russo, Nicola II, è stata verificata confrontando i suoi geni mitocondriali con quelli dei parenti. Più di recente, i resti irriconoscibili di molte vittime della strage del World Trade Center sono stati identificati per mezzo di loro geni mitocondriali. La ragione per cui i geni mitocondriali sono così utili è vincolata, in parte, al fatto che ogni mitocondrio contiene da 5 a 10 copie dei suoi geni. Poiché, di solito, ci sono migliaia di mitocondri in ogni cellula, ci sono molte migliaia di copie degli stessi geni mitocondriali in ogni cellula, mentre nel nucleo ci sono solo due copie dei geni cromosomici8. Di conseguenza, è raro non essere in grado di estrarre geni mitocondriali. Una volta estratti, il fatto che noi tutti condividiamo gli stessi geni mitocondriali con le nostre madri (e per tutta la linea materna in ascendenza e discendenza) significa che di solito è possibile confermare o rifiutare le evidenze postulate9.
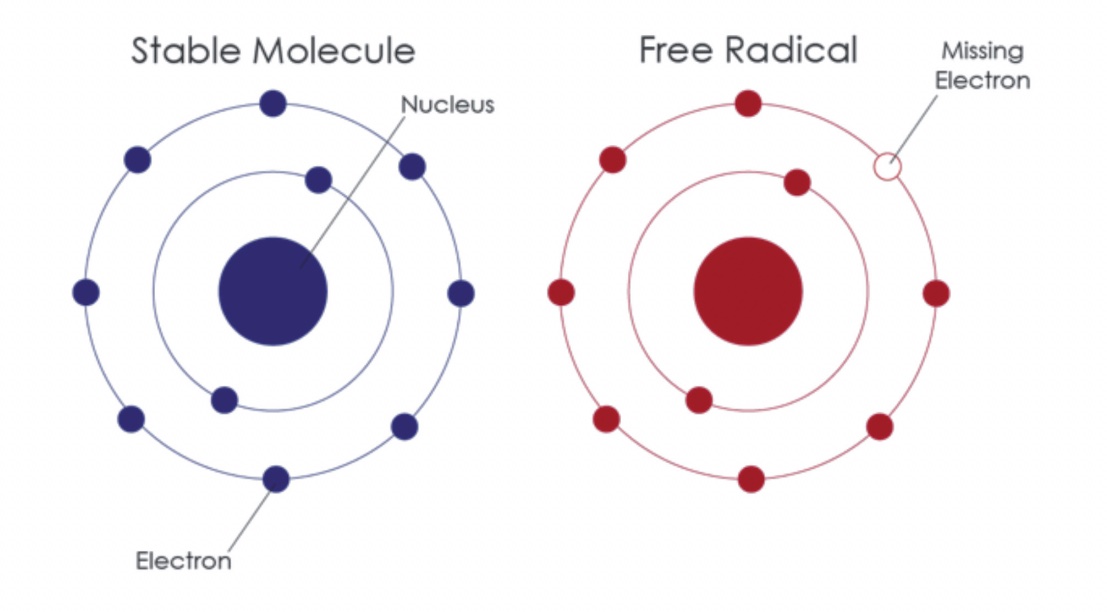
Poi c’è la “teoria mitocondriale dell’invecchiamento”10, che sostiene che l’invecchiamento umano e molte delle malattie che lo accompagnano sono causate da molecole reattive, i radicali liberi che fuoriescono dai mitocondri durante la normale funzione di respirazione cellulare. I mitocondri non sono completamente a prova di “scintilla”, ossia mentre bruciano cibo, utilizzando l’ossigeno, le “scintille” di radicali liberi permeano e danneggiano le strutture adiacenti, tra cui i geni mitocondriali stessi e geni più distanti nel nucleo cellulare. I geni nelle nostre cellule sono attaccati dai radicali liberi da 10.000 a 100.000 volte al giorno, praticamente una intromissione ogni secondo11. Gran parte del danno è riparato senza molto trambusto ma attacchi occasionali causano mutazioni irreversibili con alterazioni durevoli in sequenza genica che possono accumularsi nel corso della vita. Le cellule più seriamente compromesse muoiono, e lo spreco costante favorisce sia l’invecchiamento che le malattie degenerative. Anche molte condizioni ereditarie estreme sono legate a mutazioni causate dai radicali liberi che attaccano i geni mitocondriali. Queste malattie hanno spesso modelli di ereditarietà bizzarri e fluttuano di generazione in generazione ma, in generale, progrediscono, inesorabilmente, con l’età. Le malattie mitocondriali, tipicamente, colpiscono tessuti metabolicamente attivi come la muscolatura e il cervello, producendo convulsioni, alcuni disordini della motricità, cecità, sordità e degenerazione muscolare12.
Ad altri, i geni mitocondriali sono familiari come nel trattamento, controverso, della fertilità assistita, in cui i mitocondri sono presi da una cellula uovo (ovocita) di una donatrice e trasferiti, insieme con lo sperma, nella cellula uovo di una donna sterile13 – una tecnica nota come “trasferimento ooplasmico”14. Quando la questione diventò pubblica, un giornale britannico pubblicò l’articolo con il titolo “Bambini nati con due madri e un padre”. Questo prodotto, tipicamente vivido della stampa, non è del tutto sbagliato – mentre tutti i geni cromosomici nucleari materni provenivano dalla madre “reale”, alcuni dei geni mitocondriali provenivano dalla “madre” donatrice”, in quanto i bambini avevano effettivamente ricevuto alcuni geni da due madri diverse. Nonostante la nascita di più di 30 bambini, apparentemente sani, mediante questa tecnica, sia le preoccupazioni etiche che pratiche la misero al bando sia in Gran Bretagna che negli Stati Uniti15.
I mitocondri nella letteratura scientifica
Tutti questi aspetti riguardo i mitocondri sono familiari a molte persone attraverso i giornali e la cultura popolare. Altri lati dei mitocondri sono diventati ben noti tra gli scienziati nel corso degli ultimi due decenni, ma questi aspetti sono forse meno noti al grande pubblico. Uno dei più importanti è l’apoptosi, o morte cellulare programmata16, in cui singole cellule si “suicidano” per un bene maggiore: la salute del corpo nel suo complesso17. Da circa la metà degli anni 1990, i ricercatori hanno scoperto che l’apoptosi non è governata dai geni nucleari, come era stato precedentemente ipotizzato, ma dai geni mitocondriali. Le implicazioni sono importanti nella ricerca medica perché il fallimento nell’eseguire l’apoptosi, quando le cellule sono chiamate a farlo, è una causa principale del cancro18. Piuttosto che prendere di mira i geni nucleari, molti ricercatori stanno ora cercando di manipolare quelli mitocondriali ma le implicazioni sono profonde. Nel cancro, le singole cellule competono per la loro autonomia, sottintendendo le catene di responsabilità per l’organismo nel suo complesso. In termini dell’evoluzione iniziale, tali catene devono essere state difficili da imporre: perché cellule potenzialmente libere di vivere, avrebbero accettato una morte programmata per il privilegio di vivere in una più ampia comunità di cellule, quando ancora potevano mantenere una autonomia cellulare? In effetti, senza la morte cellulare programmata, i legami che legano le cellule in organismi multicellulari complessi avrebbero potuto non essersi mai evoluti. E poiché la morte programmata delle cellule dipende dai geni mitocondriali, può essere che gli organismi multicellulari potrebbero non essere esistiti senza i mitocondri19.
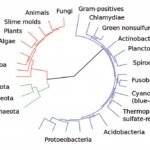
Un altro campo in cui i mitocondri sono molto visibili, oggi, è nel discorso sull’origine delle cellule eucariote20, cioè quelle cellule complesse che hanno un nucleo, da cui sono costruite tutte le piante, gli animali, le alghe e i funghi. La parola eucariote deriva dalla parola greca per “vero nucleo” la quale fa riferimento ad una sede dei geni nelle cellule. Ma la questione del nome è, francamente, insignificante. Infatti, le cellule eucariotiche contengono molti altri organuli citoplasmatici, oltre al nucleo, in particolare, i mitocondri21. Come queste prime cellule complesse si evolsero è un tema controverso. Il discorso ortodosso, al riguardo, propone che esse si sono evolute, passo dopo passo, finché un giorno una cellula eucariotica primitiva inglobò un batterio, il quale, dopo generazioni di asservimento, è diventato totalmente dipendente evolvendosi nei mitocondri. La teoria prevede che alcune remote unicellulari eucariote che non possedevano mitocondri sarebbero state le antenate di tutti noi, reliquie di un passato in cui determinati batteri si inglobarono e messi in opera come mitocondri. Ma ora, dopo anni di un’attenta analisi genetica, sembra che tutte le cellule eucariote note hanno e una volta ebbero (e poi forse persero) mitocondri22. L’implicazione è che l’origine dell’evoluzione delle cellule complesse è inseparabile dall’origine dei mitocondri: i due eventi sarebbero stati una sola e medesima cosa. Se questo è vero, allora, non solo l’evoluzione degli organismi multicellulari avrebbe richiesto mitocondri ma, così anche, l’origine stessa delle cellule eucariote. E se questo è plausibile, allora, la vita sulla terra non si sarebbe evoluta al di là dei batteri se non fosse stato per questi organuli citoplasmatici evoluti da batteri primordiali: i mitocondri23.
Un altro aspetto più nascosto, nel discorso circa i mitocondri, riguarda le differenze tra i due sessi24. Il sesso è un “rebus” ben noto: la riproduzione sessuale richiede due generi, maschile e femminile, per la riproduzione, generalmente, di un solo embrione. Una risposta al perché di questa apparente limitazione o rebus, presentata alla fine del 1970 e ora ampiamente accettata dagli scienziati, anche se relativamente poco conosciuta tra il grande pubblico, si riferisce ai mitocondri. Abbiamo bisogno di avere due sessi, perché un sesso deve specializzarsi a trasmettere i mitocondri nella cellula uovo, mentre l’altro deve specializzarsi a non trasmettere i suoi mitocondri nello sperma25. In effetti, mentre le cellule uovo contengono circa 100.000 molecole di mt DNA, quelle spermatiche ne contengono solo 1.000-1.500, destinate, sembra, a perdersi subito durante l’embriogenesi, per cui l’embrione che si svilupperà (maschio o femmina) conterrà mitocondri derivanti esclusivamente dall’ovulo cioè dal patrimonio mitocondriale materno. La costruzione di alberi filogenetici basati su dati inerenti l’mt DNA risulta più semplice e meno confusa di quella basata sul DNA nucleare in quanto non c’è ricombinazione genetica, cioè mescolanza tra geni materni e paterni26.
Il rebus dei mitocondri rimane
Tutte queste modalità di ricerca e di ragionamenti hanno ricollocato i mitocondri in una posizione che non avevano goduto dal periodo di massimo splendore durante gli anni 50 del secolo scorso, quando è stato stabilito, per la prima volta, che i mitocondri sono la sede dell’“energia” (POWER) nelle cellule, generandola quasi tutta. La rivista Science riconobbe i mitocondri nel 1999, quando dedicò la sua copertina e una sezione considerevole della rivista a questi organuli citoplasmatici sotto il titolo “Mitochondria Make A Comeback”27. [I mitocondri alla rimonta]. C’erano state due ragioni principali per questo abbandono. Uno era che la bioenergetica – lo studio della produzione di energia nei mitocondri – era stato considerato come un campo difficile e oscuro. Il secondo era relativo alla supremazia della ricerca genetica molecolare nucleare nella seconda metà del XX secolo. I biologi molecolari ignorarono i mitocondri perché non realizzavano, immediatamente, le implicazioni di vasta portata e le applicazioni della scoperta dei geni mitocondriali. Ci è voluto tempo per accumulare una banca dati, con la portata di contenuto necessaria, per affrontare molte domande impegnative relative all’antropologia, alla biogenesi, alla malattia, all’evoluzione, e altro28.
Nonostante la loro celebrità ritrovata, i mitocondri rimangono un rebus. Molte domande evolutive profonde sono a malapena poste, per non dire che non sono regolarmente discusse nelle riviste e i diversi campi di ricerca che sono cresciuti intorno ai mitocondri tendono ad essere pragmaticamente isolati nella propria competenza. Ad esempio, i meccanismi con cui i mitocondri generano energia, pompando protoni attraverso una membrana (chemiosmosis), si trova in tutte le forme di vita, inclusi i batteri più primitivi29. È un modo bizzarro di fare le cose. Secondo Nick Lane, era dai tempi di Darwin che la biologia non aveva messo a punto un’idea talmente contro l’intuizione come quelle paragonabili a quelle di Einstein, Heisenberg e Schrödinger nella fisica. Questa idea, tuttavia, si è rivelata vera e ha valso a Peter Mitchell un Premio Nobel nel 197830. Ma la domanda raramente si pone: perché un mezzo talmente peculiare di produrre energia è diventato così centrale per tante diverse forme di vita? La risposta mette in luce la questione circa l’origine della vita stessa31.
Un’altra interessante domanda, raramente affrontata, è l’esistenza di geni mitocondriali. Articoli accademici tracciano i nostri antenati risalendo all’Eva Mitocondriale e persino utilizzano i geni mitocondriali per ricostruire le relazioni tra specie diverse ma, raramente, ci si chiede il motivo per cui i mitocondri esistano. Ci sono solo assunti circa essere una reliquia di discendenza batterica. Forse, la questione è, anche, che i geni mitocondriali possono essere facilmente trasferiti in blocco al nucleo. Diverse specie hanno trasferito geni diversi al nucleo, ma tutte le specie con i mitocondri hanno anche mantenuto esattamente lo stesso gruppo di geni mitocondriali. Che cosa hanno questi geni di così speciale? La migliore risposta, come vedremo, aiuta a spiegare perché i batteri non raggiunsero mai la complessità delle cellule eucariote32. Ciò spiega perché se ciò che chiamiamo vita fosse altrove nell’universo, essa sarà probabilmente bloccata in una corsia batterica.
Ci sono molte altre simili domande, poste da pensatori percettivi nella letteratura specializzata ma raramente turbano un pubblico più ampio. Tuttavia quando poste assieme, le risposte conferiscono un racconto ininterrotto di tutta la traiettoria dell’evoluzione, dall’origine della vita stessa alla genesi delle cellule complesse e degli organismi multicellulari, al raggiungimento di organismi di maggiori dimensioni, ai sessi, all’omeotermia, al declino della vecchiaia e alla morte. Il vasto quadro che emerge ci offre nuove intuizioni riguardo il perché siamo capitati qui, e se siamo soli nell’universo, al perché abbiamo il nostro senso di individualità, al perché facciamo “sesso”, a dove facciamo risalire le nostre radici ancestrali e riguardo al perché dobbiamo invecchiare e morire – in breve, riguardo il significato della vita stessa. Lo storico Felipe Fernández-Armesto33 ha scritto: “Le storie aiutano a spiegare se stesse; se si sa come è successo qualcosa, si comincia a vedere perché è successo”, così, il “come” e il “perché” sono intimamente abbracciati quando ricostruiamo la storia della vita.
I mitocondri e l’evoluzione della vita
Quest’argomentazione sui mitocondri non è una dissertazione né specialistica né per specializzati. Essa è stata intrapresa perché le implicazioni delle ultime ricerche in materia sembrano riguardare intuizioni circa la questione del senso della vita. Quando si tratta di eventi accaduti nel passato remoto, forse miliardi di anni fa, raramente è possibile trovare risposte definitive. Tuttavia, è possibile usare quello che sappiamo, o presupponiamo di sapere, per restringere la lista delle possibilità. Ci sono indizi sparsi, a volte nei luoghi più inaspettati, e sono questi indizi che richiedono la familiarità con la biologia molecolare moderna, da qui la necessaria complessità di alcuni passaggi.
Prima di proseguire, è forse pertinente fare un accenno di biologia cellulare34 per coloro che non hanno esperienza in biologia. La cellula vivente può essere metaforicamente vista come un universo in miniatura, la forma più semplice di ciò che, dal punto di vista biologico, denominiamo vita, capace di esistenza indipendente o meno e come tale unità di base della biologia. Lo studio delle cellule è la citologia (Scienza che studia la struttura e le funzioni delle cellule animali e vegetali) e come sappiamo gli organismi si distinguono in unicellulari e pluricellulari.
Lo studio di procarioti (batteri) e eucarioti (cellule complesse) ai virus, organismi parassiti obbligati che si replicano all’interno delle cellule viventi di altri organismi, ci ha consentito una disamina dell’evoluzione della vita.
Da quando, nel 1953, James Watson e Francis Crick scoprirono la struttura del DNA, l’acido desossiribonucleico e il suo meccanismo di replicazione, si apri la strada alla conoscenza di tutto il patrimonio genetico, al suo funzionamento, alla sua decodificazione e alla possibilità di sostituire geni difettosi35. Le informazioni codificate nel DNA sono il “dogma centrale” di tutta la biologia: i geni codificano le informazioni necessarie per la formazione delle proteine e pertanto il funzionamento di ogni organismo. Il lungo doppio nastro del DNA è una sequenza apparentemente infinita di solo quattro “lettere” A, T, C, G, sinonimi dei nucleotidi che si interfacciano in coppia: adenina-timina (A-T) e citosina-guanina (C-G), così come tutte le nostre parole sono una elaborazione delle lettere alfabetiche che conosciamo. Nel DNA, la sequenza di “lettere” stabilisce la struttura delle proteine e il genoma completo è la “biblioteca” dei geni posseduti da un organismo che può arrivare a contenere miliardi di “lettere”. Un gene è essenzialmente il codice per una singola proteina che di solito richiede migliaia di lettere. Ogni proteina36 è una stringa di sub-unità chiamate amminoacidi, e l’ordine preciso di questi detta le proprietà funzionali della particolare proteina. Se la sequenza di lettere cambia a causa di una mutazione nel DNA nucleare ciò può modificare la struttura della proteina provocando, tecnicamente, una degenerazione nel codice e una degenerazione funzionale della proteina corrispondente che sarà modificata.
In breve, una determinata sequenza nucleotidica del DNA corrisponde ad una determinata frequenza degli amminoacidi, i mattoncini delle catene polipeptidiche proteiche. Come l’informazione passa dal nucleo al citoplasma il gruppo di Crick propose che da un filamento del DNA (e quindi con i relativi geni) per copia complementare venisse prodotta una molecola di RNA (acido ribonucleico). L’RNA messaggero detto mRNA si sposta dal nucleo al citoplasma cellulare dove, a livello dei ribosomi, avviene la trascrizione, ossia la sintesi proteica. La molecola che riconosce il messaggio genetico dell’mRNA, nota come RNA transfer o tRNA, è in grado di riconoscere le specifiche sequenze nucleotidiche e legare gli amminoacidi corrispondenti. In effetti, l’tRNA agisce da intermediario fra l’informazione di una sequenza nucleotidica dell’mRNA e la sequenza amminoacidica di una proteina. Ovviamente, si è accennato molto brevemente a quello che sono i processi basilari di trascrizione e traduzione per cui l’informazione genetica passa dal DNA cellulare all’RNA e poi alle catene polipeptidiche.
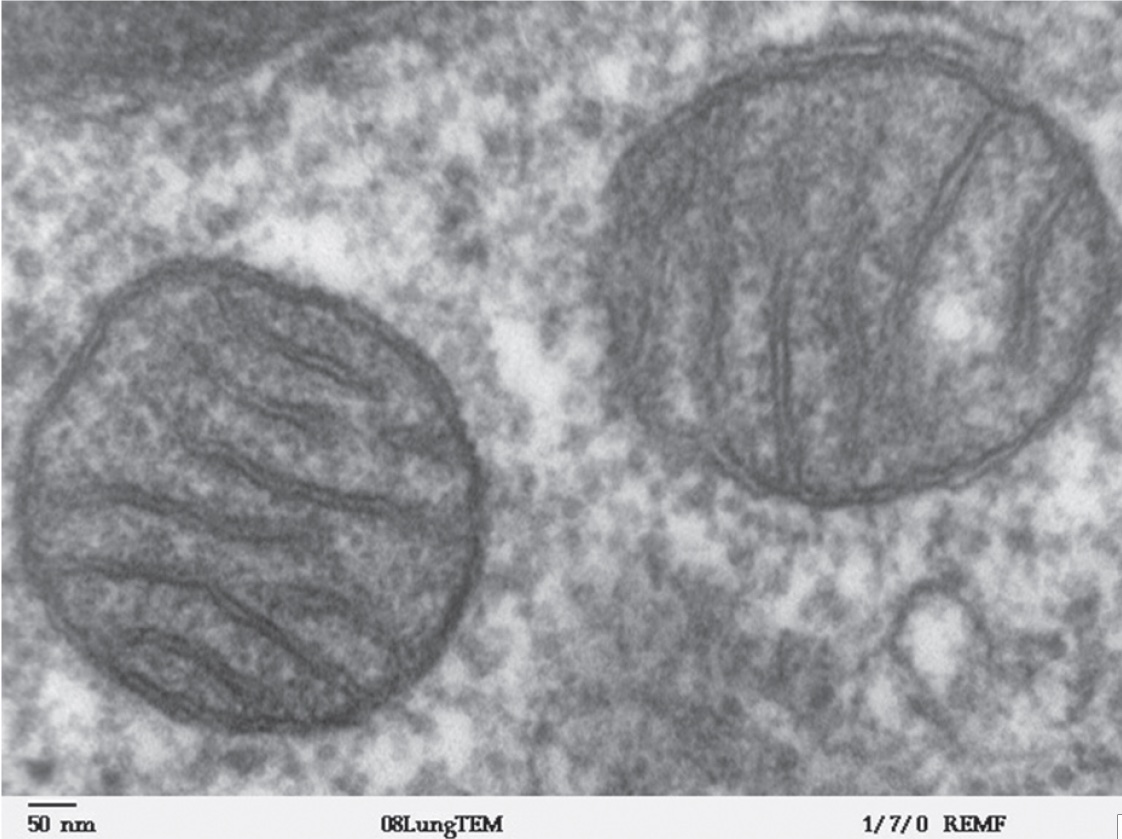
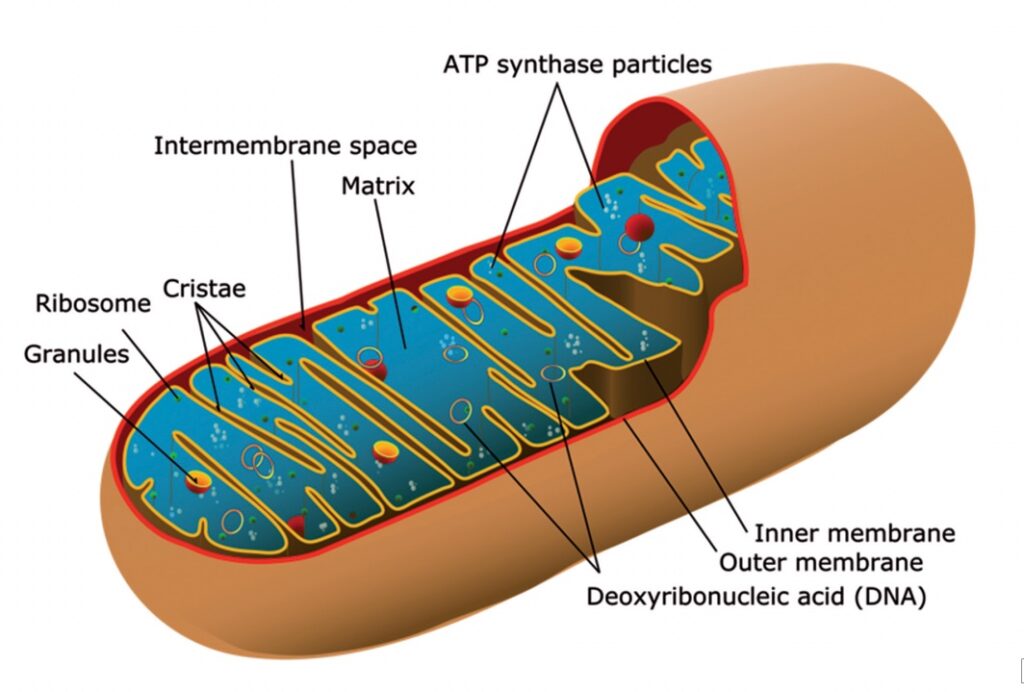
Armati di questa biologia cellulare di base, torniamo ai mitocondri. Essi sono organelli nel citoplasma cellulare dedicati ad un compito specifico: la produzione di energia. È stato detto che i mitocondri furono una volta batteri e, in apparenza, ancora un po’ li richiamano. Tipicamente raffigurati come a forma di salsicciotti o vermiformi, sono in grado di assumere forme contorte, tra cui quella di cavatappi. Le dimensioni sono di alcuni millesimi di millimetro di lunghezza (da 1 a 4 μm)37 e circa 1/2 μm di diametro. Le cellule del nostro corpo, in genere, contengono numerosi mitocondri, il numero esatto varia a seconda della richiesta metabolica di quella particolare cellula. Cellule metabolicamente attive, come quelle del fegato, dei reni, dei muscoli e del cervello, hanno centinaia o migliaia di mitocondri, che costituiscono circa il 40% del citoplasma. La cellula uovo o ovocita trasporta circa 100.000 mitocondri, mentre lo sperma ne porta solo alcune centinaia o anche meno. Ancora, ad esempio, le cellule del sangue e le cellule della pelle ne contengono molto pochi, o sono del tutto assenti. Tutto sommato, si dice che ci siano 10 milioni di miliardi di mitocondri in un essere umano adulto, che insieme costituiscono circa il 10% del nostro peso corporeo38.
I mitocondri citoplasmatici hanno due membrane, l’esterna è liscia e continua e l’interna convoluta in pieghe stravaganti o tubuli, chiamati creste39. I mitocondri, solitamente, si muovono (nuotano) nel citosol per spostarsi dove sono più necessari e, spesso, lo fanno vigorosamente. I mitocondri sono stati rilevati per prima volta nella cellula, utilizzando la microscopia ottica, come granuli, barre e filamenti, ma la loro provenienza è stata controversa fin dall’inizio. Tra i primi a riconoscere la loro importanza fu Richard Altmann, che sosteneva che i minuscoli granuli erano in realtà particelle fondamentali alla vita e, di conseguenza, le chiamò bioblasts nel 1886. Per Altmann, i bioblasts erano componenti viventi nella cellula e sosteneva che essi erano poco più di una comunità fortificata (una sorta di simbiosi) che viveva, in una sorta di simbiosi, con la cellula ospite per una reciproca protezione naturale, come i soggetti di una fortificazione dell’età del ferro. Altre strutture, come la membrana cellulare e il nucleo, sarebbero stati formati dalla comunità di bioblasts per i propri scopi, mentre il citosol (la parte “acquosa del” citoplasma) è proprio questo: un serbatoio di nutrienti racchiusi nella “rocca” microscopica40.
Le idee di Altmann non hanno mai preso piede e sono state ridicolizzate da alcuni. Altri hanno affermato che i bioblasts erano un frutto della sua immaginazione: semplicemente artefatti della sua elaborata preparazione per lavorare al microscopio. Queste dispute sono state aggravate dal fatto che i citologi erano estasiati dalla “danza” maestosa dei cromosomi durante la divisione cellulare. Per visualizzare questa “danza”, i componenti trasparenti della cellula dovevano essere colorati utilizzando coloranti. Come è accaduto, le sostanze che erano più in grado di colorare i cromosomi erano acide. Purtroppo, queste sostanze coloranti tendevano a sciogliere i mitocondri. La ossessione dei citologi per il nucleo ha fatto sì che ciò che stavano facendo era semplicemente dissolvere le prove sui mitocondri. Altre sostanze coloranti erano ambivalenti, colorando i mitocondri solo transitoriamente perché i mitocondri stessi rendevano la sostanza incolore. Il loro aspetto, piuttosto spettrale, e la loro rapida scomparsa negli studi erano poco favorevoli a sostenere una ferma convinzione della loro esistenza. Infine, Carl Benda ha dimostrato, nel 1897, che i mitocondri hanno un’esistenza corporea nelle cellule. Egli li ha definiti come “granuli, aste, o filamenti nel citoplasma di quasi tutte le cellule … che sono distrutti da acidi o solventi grassi.” Il suo termine, mitocondri è derivato dal greco mitos, che significa filo, e condrina, che significa grano piccolo.
Mentre la reale esistenza dei mitocondri era stata finalmente accettata, la loro funzione era rimasta sconosciuta. Pochi attribuivano ad essi le proprietà elementari di costruttori della vita che Altmann aveva postulato e un ruolo più circoscritto era ricercato. Alcuni consideravano che i mitocondri fossero il centro proteico o della sintesi dei grassi; altri pensavano che essi fossero la residenza di geni. Finalmente si stabilì che le sostanze coloranti venivano rese incolori dall’azione ossidante dei mitocondri stessi: un processo analogo all’ossidazione degli alimenti avviato dalla respirazione cellulare. Nel 1912, B. F. Kingbury propose che i mitocondri potessero essere i centri respiratori della cellula. Il suo suggerimento è stato dimostrato essere corretto solo nel 1949, quando Eugene Kennedy e Albert Lehninger dimostrarono che gli enzimi respiratori erano infatti situati nei mitocondri.
Anche se l’idea di Altmann circa i bioblasts era caduta in disgrazia, un certo numero di altri ricercatori ha, anche, sostenuto che i mitocondri fossero entità indipendenti legate ai batteri, simbionti che vivevano nella cellula per il reciproco vantaggio. Un simbionte è un partner in una simbiosi, un rapporto in cui entrambi i partner beneficiano in qualche modo dalla presenza dell’altro41. Rapporti reciproci possono esistere tra le cellule e i batteri, che a volte vivono all’interno di cellule più grandi come endosimbionti42. Nei primi decenni del XX secolo, quasi tutte le parti della cellula sono state considerate come possibile endosimbionti, forse modificate dalla loro reciproca convivenza, tra cui il nucleo, i mitocondri, i cloroplasti (responsabili della fotosintesi delle piante) e i centrioli (i corpi cellulari che organizzano il citoscheletro). Tutte queste teorie si basavano sull’aspetto e il comportamento, come il movimento e la divisione apparentemente autonoma e, quindi, non potevano che essere suggestive. Per di più, i loro protagonisti sono stati troppo spesso divisi da lotte circa la priorità e divisi dalla guerra e dalla lingua, raramente in accordo tra di loro43.
Gli argomenti, al riguardo, tornano alla ribalta dopo il 1918, quando lo scienziato Paul Portier pubblica il suo lavoro Les Symbiotes, affermando che: “Tutti gli esseri viventi, tutti gli organismi dall’Ameba all’uomo e dalla Crittogame ai Dicotiledoni sono costituiti da un’associazione44, l’emboîtement [l’incastro] di due organismi diversi”. Ogni cellula vivente contiene nel suo protoplasma formazioni, che gli istologi designano con il nome di mitocondri. Questi organelli, per Portier non erano altro che batteri simbionti.
Il lavoro di Portier attirò elogi e dure critiche in Francia, anche se è stato in gran parte ignorato nel mondo anglofono. Per la prima volta, tuttavia, la questione non si è centrata sulle somiglianze morfologiche tra mitocondri e batteri ma su tentativi di considerare i mitocondri all’interno della cellula. Portier ha affermato di aver fatto questo. I suoi risultati sono stati pubblicamente contestati da un gruppo di batteriologi presso l’Istituto Pasteur. Portier, comunque, non fu in grado di replicare. Egli, poi abbandonò la sua ricerca e il suo lavoro e, non essendoci stato nessuno in grado di replicarlo, è stato dimenticato.
Pochi anni dopo, nel 1925, Ivan Wallin presentò le proprie idee sulla natura batterica dei mitocondri, sostenendo che tali “simbiosi intime” erano la forza trainante dietro l’origine di nuove specie45. I suoi argomenti si sono centrati di nuovo sulla coltura dei mitocondri. Ma per la seconda volta l’interesse scemò con il fallimento di replicare il suo lavoro. Questa volta la simbiosi non è stata esclusa con tutto lo stesso veleno ma il biologo cellulare E. B. Wilson ha riassunto l’atteggiamento prevalente nella sua osservazione: “A molti, senza dubbio, tali speculazioni possono apparire troppo fantastiche per poter esserne fatta menzione nella società dei biologi, tuttavia è all’interno della gamma delle possibilità che queste speculazioni possano un giorno essere prese in seria considerazione”46.
Quel giorno si è rivelato essere mezzo secolo più tardi. Nel marzo del 1967, Lynn Margulis presentò il suo famoso paper “On the Origin of Mitosing Cells” al Journal of Theoretical Biology47, in cui resuscitava le “fantasie divertenti” delle generazioni precedenti e le ammantava con un abbigliamento di attualità scientifica. Allora l’esistenza di DNA e RNA nei mitocondri era stata dimostrata ed esempi di “ereditarietà citoplasmatica” catalogati (in cui tratti ereditati erano stati dimostrati essere indipendenti dai geni nucleari)48. Margulis si sposa poi con il cosmologo Carl Sagan e assume una visione cosmica dell’evoluzione della vita, considerando non solo la biologia ma, anche, l’evidenza geologica dell’evoluzione atmosferica e fossile di batteri e dei primi eucarioti. Lei introdusse un discernimento di consumata anatomia microbica e chimica e applicò criteri sistemici per determinare la probabilità di simbiosi. Anche così, il suo lavoro è stato respinto. Il suo paper seminale è stato respinto da 15 diversi journals prima che James Danielli, editore del Journal of Theoretical Biology, finalmente lo accettò. Una volta pubblicato, ci sono state, senza precedenti, 800 richieste di ristampa per il paper entro quell’anno. Il suo libro, “The Origin of Eukaryotic Cells”, finalmente è stato pubblicato dalla Yale University Press nel 197049. Il suo libro diventò uno dei testi di biologia più influenti del secolo. Margulis mise in un ordine talmente convincente l’evidenza che i biologi ora accettano il suo punto di vista, una volta eterodosso, come dato di fatto, almeno quando applicato ai mitocondri e cloroplasti50.
Le acri polemiche persistessero per oltre un decennio e sebbene, tuttora, molto metafisiche, erano vitali. Senza di loro l’accordo finale sarebbe stato meno sicuro. Ognuno ha accettato che ci sono effettivamente dei paralleli tra mitocondri e batteri, ma non tutti erano d’accordo su ciò che questi significassero. Certo, i geni mitocondriali sono di natura batterica, dove risiedono in un singolo cromosoma circolare (a differenza dei cromosomi “lineari” del nucleo) e sono “nudi”, ossia non avvolti in proteine istoniche51. Analogamente, la trascrizione e la traduzione del DNA in proteine è simile nei batteri come nei mitocondri. L’assemblaggio fisico delle proteine è anche gestito lungo linee simili e differisce in molti dettagli dalle funzioni standard delle cellule eucariote. Vari antibiotici funzionano bloccando l’assemblaggio delle proteine nei batteri e, anche, bloccando la sintesi proteica nei mitocondri, ma non dai geni nucleari nelle cellule eucariote.
Nel loro insieme, questi parallelismi potrebbero sembrare convincenti ma, in realtà, ci sono possibili interpretazioni alternative e sono state queste che hanno sostenuto la lunga controversia. In sostanza, le proprietà batteriche dei mitocondri potrebbero essere spiegate se la velocità di evoluzione fosse più lenta nei mitocondri rispetto al nucleo. Se così fosse, i mitocondri avrebbero più in comune con i batteri, semplicemente perché essi non si erano evoluti più velocemente. I mitocondri avrebbero mantenuto tratti più atavici. Poiché i geni mitocondriali non sono ricombinati, questa posizione era sostenibile, anche se un po’ insoddisfacente. Essa poteva essere rifiutata solo quando il tasso effettivo di evoluzione fosse conosciuto, il che a sua volta richiedeva il sequenziamento diretto dei geni mitocondriali e il confronto delle sequenze. Solo dopo che il gruppo di Fred Sanger in Cambridge ebbe sequenziato il genoma mitocondriale umano nel 1981, trasparì che il tasso di evoluzione dei geni mitocondriali era stato, addirittura, più veloce di quello dei geni nucleari. Le loro proprietà ataviche potrebbero essere spiegate solo da un rapporto diretto con i batteri e questo rapporto è stato in definitiva dimostrato di essere con un gruppo molto specifico di batteri: gli α-proteobacteria52.
Anche la Margulis non era corretta su tutto. Allineandosi con i sostenitori precedenti dell’idea di simbiosi, Margulis aveva sostenuto che un giorno sarebbe stato possibile coltivare mitocondri in coltura – secondo lei era solo una questione di trovare i giusti fattori di crescita. Oggi, sappiamo che questo non è possibile53. Il motivo di questo suo errore è stato chiarito dalla sequenza dettagliata del genoma mitocondriale: i geni mitocondriali codificano solo una manciata di proteine (13 per l’esattezza), insieme con tutti i processi genetici necessari per farli. La grande maggioranza delle proteine mitocondriali (probabilmente alcune centinaia) sono codificate da geni nel nucleo54. L’apparente indipendenza dei mitocondri è quindi falsa, e non autentica. La loro dipendenza da due genomi, mitocondriale e nucleare, è evidente anche a livello di alcune proteine che sono composte da più sub-unità, alcune delle quali sono codificati dai geni mitocondriali e altre dai geni nucleari. Perché si basano su entrambi i genomi, i mitocondri possono essere coltivati solo all’interno delle loro cellule ospiti e sono, correttamente, designati “organelli”, piuttosto che simbionti55. Tuttavia, la parola “organelli” non dà nessun suggerimento del loro passato straordinario e non offre alcuna comprensione della loro profonda influenza sull’evoluzione.56.
C’è un altro senso in cui molti biologi ancora oggi non sono d’accordo con Lynn Margulis. Questo aspetto si riferisce al potere evolutivo della simbiosi in generale. Per Margulis, la cellula eucariotica è il prodotto di molteplici unioni simbiotiche, in cui le cellule componenti sono state inglobate nell’insieme maggiore a vari livelli. La sua teoria è stata soprannominata la “serial endosymbiosis theory”, il che significa che le cellule eucariotiche sono formate da una successione di tali unioni tra le cellule, dando vita a una comunità di cellule viventi le une entro le altre. Oltre i cloroplasti e i mitocondri, Margulis cita lo scheletro delle cellule con il suo centro organizzativo, il centriolo, come il contributo di un altro tipo di batteri, le Spirochete. Infatti, secondo Margulis tutto il mondo organico è un’elaborazione di batteri collaborativi, un piccolo microcosmo come Darwin stesso aveva suggerito.
L’idea di un microcosmo è bella e stimolante ma solleva una serie di difficoltà. La cooperazione non è un’alternativa alla concorrenza. Una collaborazione tra i diversi batteri per formare nuove cellule e organismi meramente esercita una maggior sfida per la concorrenza, sfida che ora è tra gli organismi più complessi, piuttosto che tra sub-unità collaborative – molte delle quali, tra cui i mitocondri, risultano avere conservato abbastanza propri interessi. Ma la più grande difficoltà con una visione onnicomprensiva della simbiosi sono i mitocondri stessi, che agitano delle “dita”, metaforicamente parlando, in modo ammonitorio, alla potenza della collaborazione microscopica. Sembra che tutte le cellule eucariotiche hanno, o una volta ebbero (e poi forse persero) mitocondri. In altre parole, il possesso dei mitocondri è una condizione sine qua non della condizione eucariotica57.
Se la collaborazione con batteri fosse così comune potremmo aspettare di trovar tutti i tipi distinti di cellule eucariote ognuna composta da diversi tipi di microrganismi collaborativi. Naturalmente le troviamo, come c’è una grande varietà di collaborazione tra microrganismi nei fondali marini più profondi. Luoghi inaccessibili ad altre creature. Ma la scoperta più sorprendente è la presenza tra questi microrganismi di cellule eucariote primordiali che condividevano la stessa ascendenza e che ebbero (e forse persero) particelle mitocondriali. Ovviamente questo tipo di collaborazione si è sempre più evoluto tra microrganismi e cellule eucariote specializzate. In altre parole le collaborazioni primordiali che hanno ricevuto compimento tra batteri e prime cellule eucariote sono state casuali ma necessarie alla evoluzione della vita. Malgrado la competizione tra microrganismi primordiali sia durata circa 4 miliardi di anni, la collaborazione è avvenuta tra batteri primordiali e piccolissime cellule all’alba delle eucariote. In definitiva possiamo dire con certezza che l’acquisizione di piccoli organuli (mitocondri primordiali di derivazione batterica) nel citoplasma di una cellula eucariote sia stato il momento cruciale dell’evoluzione della vita58.
La ricerca continua a scoprire nuovi habitat e relazioni. Per fare un singolo esempio, una delle scoperte più sorprendenti a cavallo del millennio era, come accennato in precedenza, l’abbondanza dei piccoli eucarioti, detti pico-eucarioti, che sono organismi eucarioti picoplanctonici con una grandezza tra gli 0,2 e i 2,0 μm, vivono tra il micro-plancton in ambienti estremi, come ad esempio, i fondali oceanici dell’antartico e i fiumi acidi, ricchi di ferro, come il Rio Tinto nel sud della Spagna (conosciuto dagli antichi Fenici come il “fiume di fuoco” a causa del suo colore rosso intenso). In generale, tali ambienti sono stati considerati il dominio di batteri resistenti ed “estremofili”, cioè l’ultimo posto dove ci si potrebbe aspettare di trovare eucarioti fragili. I pico-eucarioti hanno circa le stesse dimensioni dei batteri e preferiscono ambienti simili, e così hanno generato molto interesse come possibili organismi intermedi tra batteri ed eucarioti. Eppure, nonostante le loro piccole dimensioni ed insolita predilezione per condizioni estreme, la conoscenza riguardo questi organismi rimanda a noti gruppi di eucarioti: l’analisi genetica ha mostrato che non sfidano per niente il sistema di classificazione esistente. Sorprendentemente, questa nuova fontana zampillante di variazioni delle cellule eucariote aggiunge non più di sottogruppi di gruppi esistenti, tutti conosciuti da molti anni59.
In questi ambienti insospettabili, i luoghi stessi in cui ci si aspetterebbe di trovare un “complesso” di collaborazioni uniche, troviamo molto di più. Prendiamo la più piccola nota cellula eucariote, per esempio, l’Ostreococcus tauri. È a meno di un millesimo di millimetro (1 micron) di diametro, piuttosto più piccola della maggior parte dei batteri, ma è un eucariote perfettamente formata. Ha un nucleo con 14 cromosomi lineari, un cloroplasto – e, più notevole di tutto, diversi piccoli mitocondri. E non è tutto. L’inaspettata fontana di variazione eucariotica in condizioni estreme ha gettato forse 20 o 30 nuovi sottogruppi di cellule eucariote. Sembra che tutte abbiano, e una volta ebbero, mitocondri, nonostante le loro piccole dimensioni, stili di vita insoliti, e dintorni ostili60.
Che cosa significa tutto questo? Significa che i mitocondri non sono solo giocatori in collaborazione: sono soprattutto la chiave per l’evoluzione della complessità. Se i mitocondri hanno fatto una tale differenza, in assoluto, e per la nostra vita, in particolare, vale la pena occuparsene, anche in successive argomentazioni, se come postulano alcuni ricercatori essi sono i “controllori” reconditi di molte funzioni: dalla produzione di energia, all’accumulo di calcio, alla regolazione di alcune vie metaboliche che avvengono nel citoplasma (sintesi ormonale, gluconeogenesi, sintesi proteica, attività monoaminoossidasica, produzione di calore, etc.), e, ancora, dei segnali sessuali e anche del suicido delle cellule stesse.
- P Scheffer, I. E. A century of mitochondrial research: achievements and perspectives. Mitochondrion 1: 3-31; 2000.
- Blackstone, N. W., and Green, D. R. The evolution of the mechanism of cell suicide. Bioessays 21: 84-88; 1999.
- Lane, Nick. Power, Sex, Suicide. Mitochondria and the Meaning of Life. Oxford University Press. New York, 2005.
- Ibidem, p. 3.
- Ibidem
- Scheffer, I. op. cit.
- Attardi, G. The elucidation of the human mitochondrial genome: A historical perspective. Bioessays 5: 34-39; 1986.
- Scheffer, I. op. cit.
- Attardi, G. op. cit.
- Schatz, G. The tragic matter. FEBS (Federation of European Biochemical Societies) Letters 536: 1-2; 2003.
- Barja, G. Free radicals and aging. Trends in Neurosciences 27: 595-600; 2004.
- Harman, D. The biological clock: The mitochondria? Journal of the America Geriatrics Society 20: 145-147; 1972.
- Barrit, J. A., Brenner, C. A., Malter, H. E., and Cohen, J. Mitochondria in human offspring derived from ooplasmic transplantation. Human Reproduction 16: 513-516; 2001.
- In sostanza, il trasferimento ooplasmico funziona così: i ricercatori prendono un ovulo di una donna non fertile, uno di una donna donatrice e lo sperma che deve fecondare l’ovulo modificato. Aspirano una piccola parte dell’ovulo della donatrice, cioè il citoplasma, usando un microscopio ago manovrato da un piccolo braccio robotico. Poi, questo citoplasma viene iniettato nell’ovulo della donna non fertile insieme allo sperma che deve fecondarlo.
- Lane, N. op. cit. p. 4.
- Huettenbrenner, S., Maier, S., Leisser, C., Polgar, D., Strasser, S., Grush, M., and Krupitza, G. The evolution of cell death programs as prerequisites of multicellularity. Mutation Research 543: 235-249; 2003.
- Kerr, J. F. History of the events leading to the the formulation of the apoptosis concept. Toxicology, 181-182: 471-472; 2002.
- Kerr, J. F., Wyllie A. H., and Currie A. R. Apoptosis: A basic biological phenomenon with wide-ranging implications in tissue kinetics. British Journal of Cancer 26: 239-257; 1972.
- Huettenbrenner, S., Maier, S., Leisser, C., Polgar, D., Strasser, S., Grush, M., and Krupitza, G. op. cit.
- Baldauf, S. L. The deep roots of eukaryotes. Science 300: 1703-1706; 2003.
- Vellai, T., and Vida, G. op. cit.
- Sagan, L. On the origin of mitosing cells. Journal of Theoretical Biology 14: 225-274; 1967.
- Martin, W., Hoffmeister, M., Rotte, C., and Henze, K. An overview of endosymbiotic models for the origins of eukaryotes; their ATP – producing organelles (mitochondria and hydrogenosomes) and their heterotrophic lifestyle. Biological Chemistry 382: 1521-1539; 2001.
- Whitfield, J. Everything you wanted to know about sexes. PLoS (Public Library of Science) Biology 2: 0718-0721; 2004.
- Charlesworth, B. The evolution of chromosomal sex determination. Novartis Foundation Symposium 244: 207-224; 2002.
- Da Antropologia molecolare di David Caramelli.
- Kibertis, P. A. Mitochondria make a comeback. Science 283: 1475; 1999.
- Lane, N. op. cit. p. 7.
- Harold, F. M. The 1978 Nobel Prize in Chemistry. Science 202: 1174-1176; 1978 & Chappell, J. B. Nobel Prize: Chemistry. Trends in Biochemical Sciences 4: N3-N4; 19.
- Peter Dennis Mitchell, biochimico vincitore del Premio Nobel per la chimica nel 1978 per la sua scoperta del processo chemiosmotico per la sintesi dell’ATP.
- Mitchell, P. David Keilin’s Respiratory Chain Concept and its Chemiosmotic Consequences. In Nobel Lectures in Chenistry 1971-1980.
- Lane, N. op. cit. p. 7.
- Fernández-Armesto, F. Humankind: A Brief History. Oxford University Press, 2004.
- Fruton, J. Proteins, Enzymes, Genes: The Interplay of Chemistry and Biology. Yale University Press. New Haven, USA, 1999.
- Ibidem
- Ibidem
- Il micrometro (simbolo: μm) è un’unità di misura della lunghezza corrispondente a un milionesimo di metro (cioè millesimo di millimetro).
- Lane, N. op. cit. p.11.
- Griparic, L. and van der Bliek, A. M. The many shapes of mitochondrial membranes. Traffic 2: 235-244; 2001
- Lane, N. op. cit. p. pp.11-12.
- Sapp, Jan. Evolution by Association: A History of Symbiosis. Oxford University Press, Oxford, UK, 1984.
- Dyall, S.D., Brown, M. T., and Johnson, P. J. Ancient invasions: From endosymbionts to organelles. Science 304: 253-257; 2004.
- Lane, N. op. cit. p. 13.
- Sapp, Jan. op. cit.
- Wallin, Ivan. Symbionticism and the Origin of Species. Bailliere, Tindall and Cox, London, 1927.
- Lane, N. op. cit. p. 14.
- Margulis, L. On the Origin of Mitosing Cells. Journal of Theoretical Biology, March, 1967.
- Lane, N. op. cit. p. 15.
- Margulis, Lynn. Origin of Eukaryotic Cells. Yale University Press. Yale, USA, 1970.
- Lane, N. op. cit. p. 15.
- Lane, N. Mitochondria: Key to complexity. In W. Martin (ed.), Origins of Mitochondria and Hydrogenosomes. Springer, Heidelberg, Germany, 2006.
- Attardi, G. The elucidation of the human mitochondrial genome: A historical perspective. Bioessays 5: 34-39; 1986.
- Lane, N. op. cit. p. 16.
- Fruton, J. op. cit.
- Attardi, G. op. cit.
- Ibidem
- Lane, N. op. cit. p. 17.
- Ibidem
- Ibidem
- Ibidem, p. 18.