



BIO – Medicina Costruzione Sociale nella Post-Modernità – Educational Papers • Anno XII • Numero 45 • Marzo 2023
L’idea tradizionale di cosa sia un corpo alla luce dell’idea della cellula come entità dinamica
Philip Ball, noto divulgatore delle cosiddette scoperte scientifiche fatte dai filoni considerati di avanguardia nella ricerca finanziata dall’establishment, pubblicò su AEON ad agosto dell’anno scorso un saggio apparso con il nome di “What on earth is a xenobot?” In tale lavoro Ball sintetizza i risultati delle ricerche di Iñaki Ruiz-Trillo e di Aurora Nedelcu sui principi e meccanismi delle transizioni evolutive alla vita multicellulare[1] e degli esperimenti di Douglas Blackiston, Emma Lederer, Sam Kriegman, Simon Garnier, Joshua Bongard e Michael Levin centrati sullo sviluppo di così chiamate “macchine viventi sintetiche”.[2] Il riassunto proposto da Ball si potrebbe rielaborare con la seguente parafrasi: più la ricerca contemporanea comprende come le cellule elaborano se stesse, con i suoi caratteri funzionali e morfologici, più inadeguata appare l’idea del blueprint genetico, cioè l’idea di una formula che sarebbe contenuta all’interno di tutte le cellule viventi e che deciderebbe come si sviluppi l’organismo e che aspetto abbia. Il corollario di una fine della nozione del blueprint genetico sarebbe, di conseguenza, che noi umani e gli organismi complessi non siamo un risultato di un comportamento univoco delle cellule. Un tale corollario risulta tanto sconcertante, se raffrontato al paradigma scientifico in materia sostenuto tuttora dal bio-potere, che spontaneamente mi ha predisposto a curare quest’argomentazione per le pagine di BIO.
Esperto nella comunicazione delle così designate “questioni scientifiche”, Ball ci introduce alla discussione riportandoci alla nozione moderna della morfogenesi con la retorica domanda premoderna sulla questione, vale a dire dove risieda la persona nell’embrione. Infatti, la morfogenesi, descritta oggi come la fase dello sviluppo embrionale successiva alla gastrulazione e corrispondente all’insorgenza della forma del corpo e dei suoi organi, una volta sembrava un processo fondato sull’immagine. Al riguardo, ancora nel Settecento alle capacità interpretative della nostra civiltà, incarnate negli studiosi dell’epoca, la morfogenesi sembrava un processo fondato sull’immagine e caratterizzato da un rapporto di somiglianza e parallelismo con l’oggetto denotato, ossia l’uomo. Per questo gli studiosi presumevano che il corpo dovesse, in qualche modo, esistere già, in forma minuscola, al momento del concepimento. Ancora all’inizio della modernità quest’era la nozione prevalente relativa al processo che adesso viene denominato morfogenesi. Di fatto, nel XVII secolo, il microscopista Nicolaas Hartsoeker illustrò questa teoria “preformazionista”[3] disegnando un omuncolo fetale infilato nella testa di un modello di spermatozoo.
L’idea preformazionista, come ci rammenta Ball, trova espressione moderna nella nozione, ancora condivisa e dominante nelle interpretazioni dell’establishment in materia, che postula che vi sia un piano corporeo o blueprint genetico univoco codificato nel nostro DNA. Sorprese, però, per le nostre convenzionate convinzioni, perché come ci ha anticipato Ball, più gli studiosi contemporanei, come Iñaki Ruiz-Trillo e Aurora Nedelcu,[4] iniziano a capire, a livello sperimentale, come le cellule configurerebbero se stesse, più inadeguata risulterebbe l’idea di un modello univoco e predeterminato contenuto all’interno di tutte le cellule viventi che deciderebbe come si svilupperebbe l’organismo e che aspetto avrebbe. Stando a questo filone di ricerca, sintetizzato con i lavori di Ruiz-Trillo e Nedelcu, ciò che le cellule seguirebbero non sarebbe un progetto o blueprint. In base alla loro attività di ricerca ritengono piuttosto che se le cellule possono essere considerate programmate, non sarebbero progettate con un piano rigido e predeterminato di cosa fare, ma con una serie di regole per guidare la loro auto-configurazione.[5] Come anticipato, l’implicazione scomoda di una tale proposizione per la spiegazione ufficiale dello sviluppo cellulare sarebbe che noi umani e altri organismi complessi non siamo il risultato univoco del comportamento delle cellule ma solo uno dei tanti possibili risultati.
Questa visione della cellula come fenomeno dinamico e creativo, che nel variare dei suoi aspetti manifesta l’intervento di forze o tendenze specifiche, anziché un insieme di proprietà stabilite e specificate, sfida l’idea tradizionale sostenuta ancora nella modernità occidentale riguardo cosa sia un corpo e cosa esso possa essere. Inoltre, come ci fa notare Ball,[6] l’idea della cellula come un’unità morfologica e funzionale dinamica aprirebbe alcune possibilità notevoli e persino sconcertanti sulle prospettive di reindirizzare la biologia verso nuove forme e strutture. Alla luce della ricerca sui principi e meccanismi delle transizioni evolutive alla vita multicellulare e degli esperimenti sullo sviluppo di macchine viventi sintetiche, la vita sembrerebbe improvvisamente, come argomenta Ball, più plastica e suscettibile di essere riconfigurata mediante ingegneria genetica.[7] In base all’analisi di Ball dei risultati della ricerca in questione, comprendere la contingenza e la malleabilità della configurazione multicellulare ci collegherebbe anche al nostro profondo passato evolutivo, quando gli organismi unicellulari avrebbero “intuito”, per la prima volta, i potenziali benefici del diventare multicellulari. Al riguardo, Ruiz-Trillo[8] afferma che la cellula possa essere, effettivamente, il fulcro dell’evoluzione, più dei geni o persino dell’organismo stesso. Aggiungendo che da questa prospettiva, lontani dall’apice dell’albero della vita, noi umani diventiamo solo una delle tante cose che le nostre cellule fossero e siano in grado di fare.[9]
Per documentare questa nuova visione della cellula come un’entità dinamica che nel variare dei suoi aspetti manifesta l’intervento di forze o tendenze specifiche, Ball[10] utilizza la ricerca sperimentale del biologo Michael Levin, della Tufts University di Medford nel Massachusetts, e dei suoi colleghi. Nell’opinione di Ball, in una delle più plateali documentazioni fino ad oggi presentate per dimostrare che le cellule siano capaci di più di quanto gli studiosi avessero immaginato precedentemente, questi ricercatori[11] avrebbero liberato delle cellule di rana dal loro normale percorso di sviluppo e avrebbero osservato che esse si organizzavano in modi nettamente diversi dalla precedente morfologia a rana. I ricercatori avrebbero fatto questo separando cellule da embrioni di rana che si stavano sviluppando in cellule della pelle e avrebbero semplicemente osservato cosa facevano le cellule libere.[12]
Come riferisce Ball,[13] coltivare le cellule, cioè farle crescere in una capsula di Petri o piatto da laboratorio dove verrebbero nutrite con i nutrienti di cui avrebbero bisogno, sarebbe, tutto sommato, una tecnologia matura. In generale, stando alle descrizioni disponibili sulla coltura di microrganismi in un predeterminato medium in condizioni di laboratorio controllate, tali cellule formerebbero una colonia in espansione man mano che si dividono. Ma sorpresa: stando agli esperimenti di Levin e colleghi,[14] le cellule in sviluppo in pelle di rana avrebbero mostrato di essere capaci di improvvisare innovazioni. Esse si sarebbero raggruppate in grumi approssimativamente sferici fino a diverse migliaia di cellule in ciascuno. Le cellule superficiali avrebbero sviluppato piccole sporgenze simili ai peli chiamate ciglia, presenti anche sulla normale pelle di rana. Le ciglia ondeggerebbero, in modo coordinato, per spingere i grappoli, attraverso la soluzione, proprio come remi. Questi grumi di cellule si sarebbero comportati come minuscoli organismi a sé stanti, sopravvivendo per una settimana o più, a volte diversi mesi, se forniti di cibo.[15] I ricercatori del gruppo di Levin avrebbero chiamato queste cellule xenobot,[16] derivato da Xenopus laevis, il nome latino della rana artigliata africana da cui erano state prelevate le cellule e un importante organismo modello negli studi di biologia evolutiva.
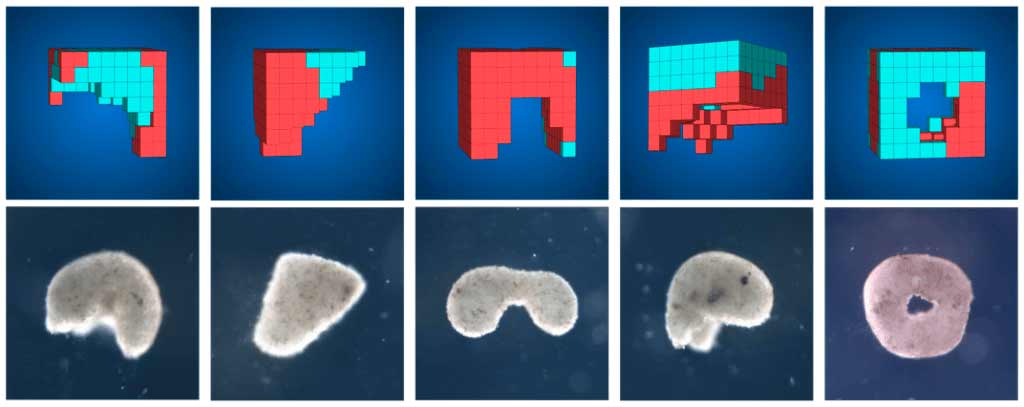
Alcuni di questi grumi di cellule non sarebbero del tutto nuovi, sostiene Ball, perché gli studiosi, in effetti, sarebbero al corrente, da più di un secolo, che un pezzo di tessuto embrionale destinato a diventare pelle, se tagliato e coltivato, farebbe crescere le ciglia. Tale pezzo di tessuto sarebbe stato chiamato “cappuccio animale” e vari studi avrebbero dimostrato che i cappucci di Xenopus[17] possono, se ricevono i giusti segnali biochimici, crescere in molti altri tipi di tessuto, inclusi neuroni, muscoli e persino tessuto di cuore pulsante.
La novità che oggi sottolinea Ball[18] sarebbe che Levin e colleghi[19] ora affermano che alcune di queste strutture o xenobot non sarebbero solo macchie casuali di cellule appiccicose ma che assomiglierebbero ad organismi autonomi. Aggiungono, inoltre, che se venissero danneggiate, le cellule guarirebbero, tornando alla forma originale.[20] In base alla loro descrizione, queste strutture potrebbero comunicare tra loro emettendo impulsi di ioni di calcio, anche se allo stato dell’arte della ricerca, Levin e i suoi colleghi[21] non avrebbero ancora un’ipotesi chiara di cosa trasmetta il messaggio. Queste strutture o xenobot si muoverebbero con uno scopo apparente, a volte circondandosi tra loro o raccogliendo in mucchi altre singole cellule attorno a loro.
Il punto interessante delle considerazioni che questi esperimenti suscitano, stando a Levin, sarebbe che questi xenobot sembrerebbero rappresentare un programma di sviluppo completamente diverso che le cellule di rana possono adottare.[22] Una spiegazione plausibile sarebbe che essendo state liberate dal loro ambiente abituale, è come se le cellule potessero scoprire un nuovo modo di vivere. La cosa sconcertante sarebbe che geneticamente non sono diverse dalle normali cellule di rana. Quindi, quest’anomalia, in relazione alle convinzioni al riguardo, seguendo il ragionamento di Ball, ci obbligherebbe a porci la domanda circa cosa codificherebbe il genoma, qualora non ci sia un piano per una rana in modo univoco.
Al riguardo, Ball[23] riferisce che sembrerebbe, invece, che i geni facciano parte di un programma molecolare che conferisce alle cellule determinate tendenze, come ad esempio a restare unite in particolari configurazioni. Le cellule sarebbero come i mattoncini Lego che possono essere assemblati in diversi modi, tranne per il fatto che le cellule si assemblano da sole. Nell’ambiente vissuto da un embrione normale, i principi dell’assemblaggio generano prima un girino e poi una rana. Ma queste non sarebbero le uniche soluzioni possibili al calcolo collettivo eseguito dalle cellule. Gli xenobot sarebbero, effettivamente, un’altra, e forse ci sarebbero altre morfologie o strutture corporee che potrebbero svilupparsi.
Stando a Ball,[24] Levin e colleghi, in particolare con la partecipazione di Josh Bongard,[25] avrebbero anche individuato un nuovo tipo di comportamento che gli xenobot potrebbero esibire. Questi ricercatori avrebbero documentato che questi pseudo-organismi possono persino replicarsi, in un certo senso. Gli xenobot posizionati in un Petri si sposterebbero per raggruppare in grumi quelle cellule sciolte che, nel corso di pochi giorni, si raggruppano organizzando nuovi xenobot che poi a loro volta si metterebbero in moto attraverso il liquido stesso, cioè il predeterminato medium di coltura.[26] Lasciati a sé stessi, gli xenobot in genere riuscirebbero a produrre solo una generazione di discendenti. I ricercatori, chiedendosi se gli xenobot avrebbero potuto fare di meglio, avrebbero effettuato simulazioni al computer alla ricerca di forme di xenobot che fossero capaci di generare più di una generazione di discendenti, utilizzando un programma di intelligenza artificiale ideato dal membro del loro team Josh Bongard.[27] Le simulazioni avrebbero suggerito che strutture come le mezze ciambelle a forma di C potrebbero spazzare via le cellule in modo più efficiente di quanto potrebbero fare gli xenobot sferoidali, creando gruppi (sferici) più grandi di “prole”.
Il team avrebbe modellato tali xenobot a mano, utilizzando minuscoli strumenti per manipolare le cellule nelle strutture a forma di C e poi lasciandole in attività su piatti di cellule fresche. Poiché la progenie risultante era più grande di quella dei precedenti xenobot, i ricercatori sarebbero stati capaci di sostenere il processo di replicazione per più generazioni. Mentre il miglior genitore sferoidale degli xenobot precedenti non poteva mai fare più di due generazioni di discendenti prima che la prole diventasse troppo piccola per riprodursi ulteriormente, i C-bot risultanti sarebbero stati in grado di generare quattro generazioni di prole, ciascuna sferoidale e di dimensioni medie gradualmente decrescenti.[28]
Questo non è, ovviamente, il modo in cui si riproducono i veri organismi viventi. Non c’è divisione cellulare coinvolta per gli xenobot e, di conseguenza, i genitori non trasmettono materiale genetico alla prole. Piuttosto, il loro comportamento assomiglierebbe più a quello che sarebbe stato osservato in alcuni robot realizzati con parti riconfigurabili, robot che avrebbero dimostrato di essere in grado di assemblare copie di sé stessi qualora venissero forniti con le parti necessarie. Stando agli esperimenti volti allo sviluppo delle cosiddette macchine viventi sintetiche condotti da Levin, Bongard e colleghi,[29] gli xenobot per replicarsi devono ricevere le parti. Questi studiosi[30]chiamano il processo auto-replicazione cinetica, vale a dire fare copie per movimento piuttosto che per riproduzione biologica. Stando a Bongard,[31] la replica sarebbe ciò che è disponibile per le entità che non possono assorbire nuova massa da sole. Tali entità possono comportarsi bene, ma non possono crescere. Tali sistemi possono solo combinare oggetti, nel loro ambiente esterno, in copie di sé stessi.[32]

Quanto lontano potrebbe l’intelligenza artificiale spingere nella generazione di entità viventi senza divisione cellulare?
Stando agli esperimenti volti allo sviluppo di macchine viventi sintetiche condotti da Levin, Bongard e colleghi,[33]combinando gli xenobot biologici con il potere esplorativo dell’intelligenza artificiale, sarebbe possibile creare una sorta di macchina vivente concepita per uno scopo. Secondo gli esperti in questo filone di ricerca, l’intelligenza artificiale potrebbe essere introdotta per esagerare una capacità innata, come afferma Bongard.[34] In base ai sui esperimenti l’intelligenza artificiale potrebbe programmare nuovi comportamenti negli organismi riorganizzando la loro morfologia piuttosto che i loro geni.[35] E questo sarebbe una svolta nell’ambito dell’assemblaggio e della replica di entità viventi senza divisione cellulare. Al riguardo, i ricercatori starebbero a chiedersi se attraverso le simulazioni si potrebbero identificare altre forme che possano assemblare strutture diverse o, forse, svolgere completamente altri compiti. In effetti, uno dei principali interessi manifestati di Bongard,[36] nel progetto in questione, sarebbe identificare quanto lontano dalla riproduzione biologia per trasmissione genica o, in questo caso, dal tipo naturale e spontaneo della replicazione degli xenobot, l’intelligenza artificiale potrebbe spingere le cose. A tale scopo ora Bongard, Levin e colleghi starebbero lavorando per incorporare diversi nuovi comportamenti negli xenobot tramite un design basato sull’intelligenza artificiale.[37]
Questa prospettiva di rielaborazione funzionale e strutturale delle cellule attraverso l’utilizzo dell’intelligenza artificiale, come ci segnala Ball,[38] comporterebbe un nuovo e controverso modo di pensare alle cellule. In questo orizzonte euristico le cellule non verrebbero pensate come blocchi di costruzione assemblati secondo un progetto o blueprint, ma come entità autonome con abilità che possono essere sfruttate per creare ogni sorta di organismi e strutture viventi. A quanto si può desumere dall’euristica[39] disponibile in materia, le cellule potrebbero essere concettualizzate come strutturate da un processo di auto-replicazione cinetica simili a robot intelligenti, riprogrammabili, muta-forma, in grado di muoversi, restare unite e di inviare segnali l’una l’altra e, in tal modo, trasformarsi in artefatti elaborati.[40]
Assumere una tale prospettiva nel modo di pensare alle cellule potrebbe anche costituire un modo più funzionale, basato sulla morfologia, che consentirebbe agli studiosi di rielaborare la concettualizzazione odierna riguardo a come i nostri corpi siano configurati durante l’embriogenesi. Come sintetizza Ball,[41] questo processo di crescita delle cellule consisterebbe in un dispiegarsi graduale e sequenziale della forma in cui ogni passaggio creerebbe le condizioni per il successivo. Fondamentalmente, questo dispiegamento, stando a quanto riportato da Ball, comporterebbe cambiamenti nello stato delle cellule stesse, innescati da segnali provenienti dall’ambiente circostante.[42] Ad esempio, quando un foglietto embrionale[43] cresce in un tessuto confinato ai suoi bordi dal resto dell’embrione, questo foglietto embrionale si troverebbe costretto a deformarsi. Le forze che agiscono sulla punta dell’uovo potrebbero quindi essere rilevate dai sensori sulle superfici cellulari che, a loro volta, attiverebbero un segnale chimico che verrebbe trasmesso ai geni. I geni verrebbero, quindi, attivati in modo da alterare le proprietà delle cellule, rendendole meno appiccicose e più mobili, tra le altre cose, e creando nuove opzioni per la forma del tessuto.[44] Da questi sviluppi teorici e dai fatti sperimentali, basati sulla ricerca di Bongard, Levin e colleghi, si desumerebbe che ci sia un dialogo continuo tra ciò che accadrebbe fuori dalla cellula e ciò che accadrebbe all’interno, tra configurazione e struttura complessiva dell’embrione in crescita e l’attività genetica, propriamente detta, all’interno dei suoi componenti.
Una talmente ricca e sottile euristica, in base alle considerazioni di Ball[45] su questi sviluppi, renderebbe difficile prevedere in quali forme crescerebbero cellule come le nostre, qualcosa resa ancora più faticosa dal fatto che, a differenza di quelle dei batteri, le nostre cellule alterano la loro attività genica in modo permanente mentre si dividono e crescono da cellule staminali embrionali a scopo generale in tessuti specializzati, disattivando alcuni geni e attivandone altri. Tuttavia, le complessità emerse in ambito sperimentale possono avviare nuove intuizioni. Infatti, come puntualizza Ball, una nuova disciplina, affianco alla biologia molecolare sintetica, chiamata morfologia sintetica, cercherebbe di accogliere e persino sfruttare questa complicazione in modo da poter sfruttare le capacità costruttive delle cellule per creare strutture e dispositivi non naturali e multicellulari completamente nuovi. Se sarà in grado di prevedere e dirigere i risultati, molto probabilmente, stando alle considerazioni di Ball[46] al riguardo, lo sforzo della morfologia sintetica si baserà sul tipo di intelligenza artificiale e altre risorse computazionali utilizzate da Levin e colleghi.
La morfologia appare scritta con leggerezza nel copione dello sviluppo evolutivo
Nelle previsioni di Ball[47] riguardo agli eventuali sviluppi di questa disciplina non sarebbe troppo inverosimile supporre che la morfologia sintetica includa la possibilità di realizzare organismi completamente nuovi. Probabilmente gli xenobot sono già un tale artefatto. Le possibili soluzioni al calcolo dell’assemblaggio delle cellule di rana, ciò che i biologi matematici chiamano stati attrattori,[48] potrebbero includere corpi microscopici totalmente diversi, simili a pesci, per esempio, o simili a vermi. Per approcciare queste probabilità riguardo a tali soluzioni, stando a Bongard, si dovrebbe utilizzare l’intelligenza artificiale in simulazioni per conoscere di più sul panorama degli attrattori di Xenopus laevis. Finora con l’intervento dell’intelligenza artificiale sarebbero stati individuati due di questi attrattori. Ciò spinge i ricercatori in questi confini ad esprimersi con entusiasmo circa le possibilità di individuarne di più anche in altre specie.
Un tale pensiero, dissacrante del paradigma dell’establishment in materia, potrebbe sfidare le nostre intuizioni su come funziona la vita, ma la verità, forse volutamente eclissata, sarebbe che gli aspetti più evidenti degli organismi, vale a dire la loro configurazione e forma complessiva, potrebbero, in realtà, essere i più superficialmente codificati, determinati non tanto da risorse genetiche pre-disegnate quanto dal modo in cui le regole dell’assemblaggio si svolgerebbero in un dato caso. In un certo senso, l’esistenza stessa di girini e rane (per non parlare degli xenobot) come prodotti vitali del genoma di Xenopus testimonierebbe a favore di questa ipotesi: i girini non sono, in effetti, morfologicamente mini-rane nel modo in cui i bambini sono mini-adulti, ma funzionano come organismi a sé stanti. Sarebbero, nei termini dell’euristica di Levin e Bongard,[49] un attrattore in cui le cellule di rana possono risiedere temporaneamente.
L’euristica (se preferite, in linguaggio epico le rivelazioni) sul potenziale morfologico delle cellule mostrerebbe, stando a Ball,[50] perché il fatto che siamo così geneticamente simili ai nostri parenti evolutivi, avendo il 99% di sovrapposizione genetica con gli scimpanzé e l’84% con i cani, coglierebbe solo in parte la questione di tale potenziale. La sua interpretazione, per spiegare meglio il potenziale morfologico delle cellule, sarebbe che la maggior parte delle nostre risorse genetiche sembrerebbero funzionalmente riservate a sostenere le capacità costruttive e adattive delle cellule che ci conformano.[51] Le forme esatte che configurano possono sembrare un’improvvisazione. Infatti, da questa prospettiva, tutto ciò che servirebbe per fare la differenza tra un corpo umano e un corpo di cane sarebbe una piccola messa a punto delle regole che presiederebbero lo sviluppo. Naturalmente, tali differenze sarebbero vitali per il successo evolutivo nella nicchia abitata da un organismo ma, anche così, stando agli studiosi alla base di quest’argomentazione, la morfologia appare scritta con leggerezza nel copione dello sviluppo.
Conforme alle interpretazioni di Ball circa l’euristica in materia[52] ciò che la genetica evolutiva evincerebbe utilizzando un suo paradigma sarebbe che la morfologia appare scritta con leggerezza nel copione dello sviluppo evolutivo. Questa sconcertante puntualizzazione, che sembrerebbe far traballare il trionfalismo delle promesse di una medicina basata su un presunto determinismo genomico metaforicamente matematico e riducibile alla volontà di un progetto a carattere eugenetico, costituisce la ragione per cui decisi di curare e presentarvi quest’argomentazione in queste pagine di BIO, editorialmente dedicate alla decostruzione sociale e concettuale della medicina.
Ma le sconcertanti considerazioni che si desumono dall’euristica odierna sullo studio delle cellule e del loro genoma[53] non si limiterebbero a suggerire una leggerezza della sua scrittura. Esse, per di più, metterebbero in evidenza che non ci sarebbe stata molta innovazione nei geni stessi da quando i primi organismi multicellulari complessi, cioè i metazoi,[54] sarebbero apparsi circa 750 milioni di anni fa.[55] Inoltre, lo shock della nostra civiltà nello scoprire, grazie proprio al Progetto Genoma, che avremmo solo un numero di geni codificanti proteine pari a quello del minuscolo verme terrestre Caenorhabditis elegans,[56] che sarebbe costituito solo da 2.000-3.000 cellule, potrebbe essere stato, in gran parte, dovuto a un senso di orgoglio ferito, come suggerisce Ball.[57] Infatti, una tale documentazione circa il numero di geni codificanti proteine mette apertamente in discussione la convinzione, anche religiosa, che noi umani siamo speciali.
Eppure questi sviluppi nell’euristica della conoscenza della nostra civiltà circa le cellule e il loro genoma piuttosto che essere stigmatizzati come controversi o sconcertanti potrebbero essere apprezzati come un’indicazione che, in primo luogo, avevamo una visione sbagliata dello scopo delle risorse genetiche, oppure come una segnalazione che ci invita a prendere atto, nelle sedi che gestiscono la validità della conoscenza, che il potenziale morfologico delle cellule non fornirebbe un piano prestabilito di sviluppo ma che, semplicemente, aiuterebbe a creare opzioni.

Dopotutto, come puntualizza Ruiz-Trillo,[58] la maggior parte dei geni e delle capacità necessarie per uno stile di vita multicellulare sarebbero stati già presenti nei nostri antenati unicellulari. Al riguardo, si potrebbe annoverare, seguendo gli sviluppi della ricerca, che essi possedevano già la capacità di inviare segnali l’un l’altro per consentire un comportamento cooperativo, restare uniti e specializzarsi in diversi tipi di cellule. Oggi tali capacità sarebbero verificabili, seguendo Ruiz-Trillo,[59] nelle amebe unicellulari come la Dictyostelium discoideum,[60] che sotto stress si riunirebbe in “corpi” multicellulari.
Ruiz-Trillo e i suoi collaboratori[61] pensano che le risorse genetiche necessarie per tale comportamento multicellulare derivino in gran parte dai meccanismi di regolazione dei geni – attivandoli e disattivandoli – piuttosto che da qualsiasi novità nei geni stessi. In base alle loro interpretazioni, gran parte dell’innovazione nel contenuto genico osservata nella transizione verso la multi-cellularità sarebbe radicata nel pervasivo armeggiare con famiglie di geni preesistenti. Questi ricercatori sarebbero arrivati a tale conclusione attraverso i loro studi sull’ameba Capsaspora owczarzaki,[62] che sarebbe uno dei parenti evolutivi più stretti dei primi organismi multicellulari. Capsaspora avrebbe più geni coinvolti nelle funzioni regolatrici di qualsiasi altro organismo unicellulare, per lo più proteine codificanti chiamate fattori di trascrizione.[63] Ruiz-Trillo e colleghi avrebbero documentato che le reti di interazione biomolecolare che queste proteine governano in Caspaspora si troverebbero spesso anche negli animali. In altre parole, le reti sarebbero già preparate e pronte a partire prima che la vera multi-cellularità fosse decollata.[64]
Stando alle interpretazioni di Ruiz-Trillo, in un certo senso, la nostra continua vicinanza al confine tra stili di vita unicellulare e multicellulare potrebbe essere vista come la ragione per cui noi, umani, (e quasi tutti i metazoi o animali che presentano una condizione monocellulare allo stadio di germe ma che raggiungono durante lo sviluppo una condizione pluricellulare con un numero più o meno elevato di elementi somatici variamente differenziati) siamo inclini al cancro, in cui le nostre cellule sembrano aver abbandonato la moderazione richiesta per la vita multicellulare e essere ritornate all’eccesso proliferante della singola cellula.[65] Nella visione sviluppata da Ruiz-Trillo e colleghi, il problema più importante degli organismi multicellulari probabilmente sarebbe il fatto che alcune cellule finiscono andando da sole. Stando a lui, infatti, molti dei geni che sarebbero cruciali per la multi-cellularità animale sarebbero geni coinvolti nella malattia del cancro.[66] Queste sue considerazioni lo portano a suggerire che forse la condizione di organismi multicellulari potrebbe essere anche interpretata come una condizione contro il modo di vivere ancestrale,[67] vale a dire riferito alla vita ancora unicellulare, e, di conseguenza, come qualcosa che richiede uno sforzo e un’energia continui per mantenersi.[68]
Il rovescio della medaglia di questa visione sarebbe che ugualmente la vita unicellulare tende a diventare collettiva. Come espressione radicale di questa tendenza delle cellule a partecipare in un insieme, Ball[69] riporta che nemmeno le cellule tumorali sarebbero inclini all’auto-replicazione, ignare delle altre cellule intorno a loro. Al riguardo Ball segnala che molti tumori assomigliano meno a masse indifferenziate di cellule che si moltiplicano selvaggiamente e più a una versione squilibrata della crescita degli organi. Stando alle sue conoscenze della questione, anche le cellule tumorali possono differenziarsi e specializzarsi, come se inseguissero una nuova traiettoria. In merito, Ball[70]aggiunge che un tumore, lungi dal crescere incurante dei tessuti ospiti che lo circondano, può integrarsi con quei tessuti e persino impossessarsene per i propri fini. In un certo senso, nella visione dei ricercatori di cui Ball si fa interprete, i tumori rappresenterebbero una morfologia alternativa delle nostre stesse cellule.
Questo stesso modello di innovazione evolutiva attraverso la novità nell’uso dei geni, piuttosto che nei geni stessi, avrebbe continuato ad agire mentre la vita multicellulare diventava più complessa. A questo riguardo, il biologo molecolare Michel Morange[71] dell’École Normale Supérieure de Paris afferma che i principali cambiamenti osservati durante l’evoluzione sarebbero più la conseguenza della riorganizzazione delle reti di regolazione genica che della modifica dei legami proteici che le formano. Nel 2011 i biologi Craig Lowe, David Haussler e i loro colleghi avrebbero individuato[72] quali tipi di cambiamenti normativi sarebbero stati coinvolti nell’evoluzione dei vertebrati dalla loro prima apparizione circa 650 milioni di anni fa. Al riguardo avrebbero confrontato i genomi di vari vertebrati – umani, mucche, topi e due tipi di pesci (spinarello e medaka, o pesce di riso giapponese) – per individuare quali sequenze genetiche condividevano e quali, presumibilmente, avrebbero avuto anche i loro antenati comuni.
I ricercatori del gruppo di Lowe e Haussler[73] avrebbero preso in considerazione parti delle sequenze solitamente non esaminate in tali raffronti filogenetici, vale a dire i cosiddetti elementi non esonici,[74] che non rientrerebbero nelle sequenze che codificano le proteine. Le sequenze non esoniche sarebbero spesso ritenute spazzatura genomica casuale accumulata per caso, ma Lowe e colleghi avrebbero ipotizzato che se alcuni elementi non esonici fossero risultati altamente conservati, vale a dire che si ripresentassero più o meno invariati in specie diverse, allora probabilmente avrebbero un ruolo funzionale nella cellula. Nell’interpretazione di Lowe e colleghi, ciò avrebbe significato che tali elementi non esonici sarebbero soggetti a pressioni selettive, che li conserverebbero, mentre ci si aspetterebbe che la spazzatura casuale degeneri rapidamente e diverga in sequenza tra specie diverse. Al riguardo, i ricercatori del gruppo di Lowe avrebbero congetturato che tali elementi non esonici conservati sarebbero probabilmente coinvolti nella regolazione dell’attività dei geni.[75]
Gli studiosi in questione avrebbero individuato che, piuttosto che cambiamenti fluidi e graduali nelle frequenze degli elementi non esonici, sembrerebbe che si siano verificate tre distinte ere di cambiamento da quando i vertebrati si sarebbero evoluti per la prima volta.[76] Nella loro interpretazione, fino a circa 300 milioni di anni fa, quando i mammiferi si sarebbero separati da uccelli e rettili, i cambiamenti nella regolazione sembrerebbero essere avvenuti principalmente in parti del genoma vicine ai fattori di trascrizione e ai geni chiave che li controllano.[77] Poi, tra 300 e 100 milioni di anni fa, quei cambiamenti si sarebbero attenuati. Invece, sarebbero state osservate modifiche vicino ai geni che codificano per le molecole proteiche che fungono da recettori del segnale sulla superficie cellulare. Nelle parole di Ball,[78] ciò che sembrava importante per questi cambiamenti evolutivi non sarebbe un cambiamento nel contenuto delle cellule, ma nel modo in cui parlerebbero tra loro, cioè il dialogo che renderebbe possibile la multi-cellularità. Infine, stando a Lowe[79] a partire da 100 milioni di anni fa, durante l’emergenza dei mammiferi placentari (cioè tutti i mammiferi tranne i marsupiali e i monotremi come l’echidna), i cambiamenti regolatori sembrerebbero essere associati a meccanismi di modifica della struttura delle proteine dopo che sarebbero state sintetizzate in forma grezza, in particolare per le proteine associate alla trasmissione di segnali all’interno delle cellule.
I cambiamenti evolutivi sarebbero vincolati al modo in cui le cellule parlerebbero tra loro
A questo punto della ricerca, stando a Ball,[80] si potrebbe quindi ritenere che l’evoluzione abbia successivamente avviato modi per innovare e generare nuovi organismi rimescolando, in primo luogo, il modo in cui i geni dello sviluppo vengono attivati e disattivati, quindi il modo in cui le cellule comunicano e, infine, il modo in cui le informazioni vengono trasmesse all’interno delle cellule. In tutti i casi, l’azione si concentrerebbe su come le cellule interagiscono e rispondono l’una con l’altra, piuttosto che su cosa fanno le singole cellule. In altre parole, sarebbero modifiche alle regole di assemblaggio multicellulare. A parer di Lowe,[81] sembra che il nostro set di circa 20.000 geni sia in grado di costruire morfologie che variano enormemente sia nella forma che nella complessità. Lui e i suoi colleghi[82] pensano che la complessità sia codificata da quante regole siano codificate nel genoma per controllare quando e dove quei geni si attivano e disattivano. Il verme C elegans, pur avendo all’incirca lo stesso numero di geni, avrebbe ben poco di questa sofisticazione normativa e, quindi, sarebbe bloccato in una forma a bassa complessità.
Nell’interpretazione persuasiva di Lowe e colleghi, il potenziale generativo delle cellule attrezzate per la costruzione multicellulare si sarebbe evidenziato quasi non appena la vita a scala multicellulare sarebbe diventata un’opzione di stile di vita in termini evolutivi.[83] Stando a loro, nell’esplosione del Cambriano,[84] circa 540 milioni di anni fa, sarebbero apparsi tutti i tipi di strane forme del corpo, molte delle quali non sarebbero più esibite da nessun organismo sulla Terra. Secondo Ball,[85] forse dovremmo considerare quelle infinite forme bellissime dimenticate, per prendere in prestito la frase risonante di Charles Darwin, come un’illustrazione del potenziale costruttivo della cellula metazoica, vale a dire un’espressione esuberante della gamma di soluzioni al problema dell’assemblaggio cellulare, che la selezione naturale, poi, rigorosamente avrebbe ridotto.
Infine, e tornando al punto centrale di questa riflessione circa le cellule come entità dinamiche e creative, riconoscere che la forma umana sia un risultato accidentale del modo in cui le nostre cellule sarebbero capitate, addirittura per semplici soluzioni creative, solleva, come a più riprese sottolineato, alcune domande scomode, come ad esempio, se potrebbe esserci un tipo di organo o tessuto che le nostre cellule potrebbero produrre ma che normalmente non ne avrebbero l’opportunità. Oppure se le nostre cellule immobili potrebbero ricordare forme corporee evolutive più antiche.
Al riguardo, Bongard propone che forse almeno alcuni degli attrattori che gli algoritmi di intelligenza artificiale potrebbero aiutarci a identificare negli assemblaggi di tipo xenobot siano echi di obiettivi passati della selezione naturale.[86] Approfondendo questa considerazione di Bongard, Ball aggiunge che almeno alcune di queste nuove configurazioni potrebbero suggerire agli studiosi della questione nuove intuizioni sugli ambienti e le pressioni selettive che avrebbero agito su questi organismi nel loro lontano passato e su come abbiano risposto evolutivamente. In un certo senso, questi attrattori sarebbero come fossili, cioè potrebbero essere intesi come scorci parziali del passato.[87]
L’euristica circa le cellule come entità dinamiche solleva anche domande su quanto i gruppi sociali portatori di interesse in tale attività potranno rimodellare le forme biologiche, inclusa la nostra forma umana. In un certo senso, sappiamo già che il corpo umano mostra una notevole plasticità. In tal senso, nessun segnale genetico dice a un embrione di dividersi in gemelli identici, per esempio. Una tale divisione sarebbe da intendersi solo come un modo in cui si verificano le regole di assemblaggio. E anche modifiche genetiche relativamente modeste alle regole potrebbero generare corporature marcatamente diverse come, ad esempio, la condizione ereditaria chiamata sindrome di Kartagener, malattia ereditaria e congenita caratterizzata da una serie di deviazioni dalla norma della posizione e della funzionalità di alcuni organi, frequenti soprattutto nel sistema cardiocircolatorio. Brevemente spiegato, sarebbe come se un passaggio chiave nella formazione iniziale del piano corporeo fosse andato storto, ma poi le cellule lo avrebbero adattato come meglio potevano. Problemi di sviluppo come la spina bifida, in cui il tubo neurale che diventerà il midollo spinale non riesce a chiudersi, avrebbero molte cause complesse e non perfettamente comprese che possono tutte portare all’esito sbagliato delle regole di assemblaggio nella morfogenesi. Sbagliato, cioè per la salute del futuro bambino.

A volte le regole si traducono in un risultato morfologico solo leggermente diverso da quello più comune: un dito in più, un arto più corto, una statura complessiva diversa, una palatoschisi.[88] Questi sarebbero errori solo se scegliamo di renderli tali. Occasionalmente, infatti, una forma del corpo non standard costituisce un problema solo perché, come società, non la prendiamo adeguatamente in considerazione, sia fisicamente, esteticamente o socialmente. Da questa prospettiva delle cellule come entità dinamiche e creative, che nel variare dei suoi aspetti manifesta l’intervento di forze e tendenze specifiche, ci sarebbero molti modi in cui noi umani possiamo conformarci, perché il risultato non starebbe scritto inflessibilmente nel nostro genoma. In questa visione di apertura, che intende solo allargare le nostre idee sociali sulla vita, ciò che le cellule ottengono sarebbero soltanto linee guida per l’assemblaggio e il risultato non sarebbe prescritto, forse perché, nell’interpretazione dei ricercatori accennati in quest’argomentazione, la natura avrebbe intuito che ci sono modi migliori e più versatili per le soluzioni morfologiche da quello della prescrizione.
______________Note _________________
[1] Ruiz-Trillo Iñaki and Nedelcu Aurora M. Editors. Evolutionary Transitions to Multicellular Life. Principles and mechanisms. Springer, 2016
[2] Douglas Blackiston, Emma Lederer, Sam Kriegman, Simon Garnier, Joshua Bongard and Michael Levin. A cellular platform for the development of synthetic living machines. Science Robotics. Vol 6, Issue 52, 17 March 2021
[3] Il preformismo, o teoria preformazionista, è la teoria, sorta nel XVII secolo, secondo la quale l’animale adulto, con tutti gli organi e i caratteri ereditari, si trova già in miniatura nel germe, ossia nell’uovo o nello spermatozoo. Al preformismo si contrapponeva la teoria dell’epigenesi, secondo la quale, al contrario, l’embrione si sviluppa, a partire da un germe indifferenziato, mediante la formazione successiva delle varie parti dell’organismo. Negli anni settanta del XVII secolo, grazie al microscopio, Francesco Redi dimostrò che anche gli insetti si riproducevano mediante uova, in seguito a queste osservazioni si affermò la dottrina dell’ovismo secondo la quale tutti gli animali, anche quelli vivipari, si sarebbero riprodotti mediante uova come gli animali ovipari. Quasi contemporaneamente, dall’osservazione degli spermatozoi nel liquido seminale fatta da Antoni van Leeuwenhoek nel 1677, nasceva la contrapposta dottrina dello spermatismo, secondo la quale l’embrione sarebbe già presente nello spermatozoo, mentre l’uovo avrebbe avuto la funzione di nutrirlo. Nel 1694 Nicolas Hartsoeker ipotizzò che l’intero feto, un “omuncolo” replica microscopica dell’essere in gestazione, alloggiasse nello spermatozoo con l’estremità cefalica nella testa dello spermatozoo. Entrambe le teorie, l’ovismo e lo spermatismo, sottolineavano l’importanza della riproduzione per lo studio degli esseri viventi, ed entrambe condividevano la teoria secondo la quale l’animale adulto si trova preformato già nelle cellule germinali. La teoria preformista fu sostenuta per la prima volta dall’olandese Jan Swammerdam il quale, nel Miraculum naturae sive uteri muliebris fabrica (1672) negò che negli insetti esistesse una vera metamorfosi: la farfalla, per es., per Swammerdam è già presente interamente, con i suoi organi già distinti, nelle uova del bruco. Secondo Swammerdam tutti i germi preesistevano dall’inizio del mondo, essendo la Creazione un atto unico. Pertanto al momento della creazione nelle ovaie di Eva si sarebbero trovati, in miniatura, tutti gli uomini destinati a nascere fino alla fine del mondo. Lo sviluppo degli esseri viventi non era altro che svolgimento delle parti impacchettate nel germe, con successive mutazioni quantitative (accrescimento e allungamento). Anche scienziati di primo piano come Leibniz, Bonnet e Spallanzani furono partigiani del preformismo, e il preformismo fu l’ipotesi giudicata più credibile dall’Encyclopédie di Diderot e d’Alembert. La polemica tra i fautori dell’epigenesi e quelli del preformismo sarà feroce per tutto il secolo XVIII e si concluderà solo nel XIX secolo con l’affermazione definitiva della teoria cellulare. Fonte: Aldo Devizzi, “Preformismo” ed “Epigenesi, Dottrina della”. In: Dagobert D. Runes (a cura di), Dizionario di Filosofia, Aldo Martello editore, Milano, 1963 / Clara Pinto Correia, The Ovary of Eve: Egg and Sperm Preformation. University of Chicago Press, Chicago, 1997
[4] Ruiz-Trillo Iñaki and Nedelcu Aurora M. op. cit. 2016
[5] Ibidem
[6] Philip Ball. What on earth is a xenobot? AEON, 30 Aug 2022
[7] Ibidem
[8] Ruiz-Trillo Iñaki and Nedelcu Aurora M. op. cit. 2016
[9] Ibidem
[10] Philip Ball. What on earth is a xenobot? AEON, 30 Aug 2022
[11] Douglas Blackiston, Emma Lederer, Sam Kriegman, Simon Garnier, Joshua Bongard and Michael Levin. op. cit. 17 March 2021
[12] Ibidem
[13] Philip Ball. What on earth is a xenobot? AEON, 30 Aug 2022
[14] Douglas Blackiston, Emma Lederer, Sam Kriegman, Simon Garnier, Joshua Bongard and Michael Levin. op. cit. 17 March 2021
[15] Ibidem
[16] Uno xenobot è un organismo semi-sintetico progettato con l’ausilio di un calcolatore affinché svolga una funzione desiderata e realizzato combinando insieme diversi tessuti biologici. Questo genere di organismi prende il nome dallo xenopo liscio, una rana acquatica endemica dell’Africa australe. Come sottolineato dai loro creatori, non essendo né robot tradizionali né nuove specie animali, gli xenobot fanno parte di una nuova classe di artefatti, oggetti artificiali che sono organismi viventi e programmabili. Gli xenobot sono larghi meno di un millimetro e sono composti esclusivamente da due cose, ossia cellule ectodermiche e miocardiotici derivati da cellule staminali ricavate da embrioni di rospo (il sopraccitato xenopo) allo stadio di blastula. Nello specifico, le cellule ectodermiche forniscono il supporto rigido, mentre i miocardiociti fungono come piccoli motori, contraendosi ed espandendosi in volume, in modo da fornire una propulsione in avanti allo xenobot. La sua forma e la distribuzione in esso dei due tipi di cellule sopraccitati sono progettati automaticamente da un software che utilizza un algoritmo evolutivo al fine di poter portare a termine un compito preciso. Grazie a tali software sono quindi stati progettati xenobot in grado di camminare, nuotare, trasportare carichi e lavorare assieme in gruppo allo scopo, ad esempio, di radunare detriti sparsi sulla superficie di una piastrina in modo da formare delle pile ordinate. Gli xenobot possono sopravvivere per settimane senza un apporto nutritivo esterno e possono persino curare sé stessi dopo aver subito una lieve lacerazione. Alla fine del novembre 2021 è stata annunciata l’osservazione di un tipo di riproduzione sviluppato autonomamente dagli xenobot e mai precedentemente osservato in natura. Il principio di base di tale riproduzione è detto replicazione cinetica, un fenomeno già noto ma solo a livello molecolare, non essendo mai stato osservato né a livello di una semplice cellula né, tantomeno, a quello di un organismo intero. Quello che è stato osservato è che gli xenobot sono in grado spostare e combinare cellule staminali disperse nel loro ambiente al fine di creare assiemi che sembrano e si muovono come loro stessi, creando in pratica, con un processo di “gestazione” che richiede pochi giorni e durante il quale le cellule assemblate “maturano”, delle copie funzionali di sé stessi, e che questa capacità non deve essere specificamente evoluta o introdotta dalla manipolazione genetica. Ciò, secondo gli autori della ricerca, dimostra che l’ingegneria dell’intelligenza artificiale può progettare cluster in grado di replicarsi nel modo migliore e di svolgere contemporaneamente un lavoro utile, e suggerisce che le tecnologie future potrebbero, con poche indicazioni esterne, diventare sempre più utili man mano che si diffondono. La realizzazione degli xenobot è stata annunciata nel gennaio 2020 da un gruppo di ricercatori divisi tra l’università del Vermont, la Tufts e l’Istituto Wyss di Harvard; da allora, essi sono stati utilizzati soprattutto come strumento scientifico utile a comprendere come le cellule cooperino tra loro per costruire corpi complessi durante la morfogenesi. Il comportamento e la biocompatibilità degli xenobot hanno fatto immaginare per loro una serie di potenziali applicazioni in cui potrebbero risultare fondamentali. Dato che gli xenobot sono composti esclusivamente da cellulare di rospo, essi sono biodegradabili e ciò, unitamente alla dimostrata capacità di poter radunare carichi microscopici in pile ordinate, ha fatto pensare a un loro utilizzo nella rimozione delle microplastiche dagli oceani: gli xenobotpotrebbero infatti realizzare, a partire da minuscoli pezzi di plastica, delle palle sufficientemente grandi da poter essere raccolte da una barca o da un drone e quindi trasportate in un centro di riciclaggio. Contrariamente alle tecnologie tradizionali gli xenobot non inquinano né durante il loro lavoro né durante la loro degradazione, essi infatti ricavano energia dal grasso e dalle proteine naturalmente localizzati nei loro tessuti, che durano circa una settimana e finiti i quali gli xenobot si trasformano semplicemente in piccoli ammassi di cellule morte. Secondo i loro creatori, in future applicazioni mediche, come la consegna mirata di medicinali, gli xenobot potrebbero essere realizzati con cellule del paziente stesso, in modo tale da evitare tutti i problemi di risposta immunologica scatenati invece da sistemi simili basati sull’uso di micro-robot. Simili xenobot troverebbero molte altre applicazioni, tra cui la rimozione di placche dalle arterie, e, con l’aggiunta di altri tipi di cellule e di ulteriore bioingegnerizzazione, essi potrebbero essere utilizzati per localizzare tumori. Fonte: Sam Kriegman, Douglas Blackiston, Michael Levin e Josh Bongard, A scalable pipeline for designing reconfigurable organisms. in Proceedings of the National Academy of Sciences, vol. 117, n. 4, 13, pp. 1853-1859, January 13 2020
[17] Lo xenopo liscio o platanna è una rana acquatica appartenente alla famiglia Pipidae, endemica dell’Africa australe. È un importante organismo modello negli studi di biologia evolutiva dello sviluppo.
[18] Philip Ball. What on earth is a xenobot? AEON, 30 Aug 2022
[19]Douglas Blackiston, Emma Lederer, Sam Kriegman, Simon Garnier, Joshua Bongard and Michael Levin. op. cit. 17 March 2021
[20] Ibidem
[21] Ibidem
[22] Ibidem
[23] Philip Ball. What on earth is a xenobot? AEON, 30 Aug 2022
[24] Ibidem
[25] Sam Kriegman, Douglas Blackiston, Michael Levin and Josh Bongard. Kinematic self-replication in reconfigurable organisms. Biophysics and Computational Biology, 118 (49) e2112672118, Nov. 29, 2021
[26] Ibidem
[27] Ibidem
[28] Ibidem
[29] Douglas Blackiston, Emma Lederer, Sam Kriegman, Simon Garnier, Joshua Bongard and Michael Levin. op. cit. 17 March 2021
[30] Sam Kriegman, Douglas Blackiston, Michael Levin and Josh Bongard. op. cit. Nov. 29, 2021
[31] Ibidem
[32] Ibidem
[33] Ibidem
[34] Ibidem
[35] Ibidem
[36] Ibidem
[37] Ibidem
[38] Philip Ball. What on earth is a xenobot? AEON, 30 Aug 2022
[39] Euristica: parte dell’epistemologia e del metodo scientifico che si occupa di favorire la ricerca di nuovi sviluppi teorici, nuove scoperte empiriche e nuove tecnologie, con un approccio alla soluzione dei problemi che non segue un chiaro percorso, ma che si affida all’intuito e allo stato temporaneo delle circostanze al fine di generare nuova conoscenza. In particolare, l’euristica di una teoria dovrebbe indicare le strade e le possibilità da approfondire nel tentativo di renderla “progressiva”, in grado cioè di prevedere fatti nuovi non noti al momento della sua elaborazione.
[40] Philip Ball. What on earth is a xenobot? AEON, 30 Aug 2022
[41] Ibidem
[42] Ibidem
[43] I foglietti embrionali, o foglietti germinativi, indicano nella biologia evolutiva degli organismi pluricellulari la prima differenziazione di un embrione
in diversi strati cellulari, dai quali successivamente si sviluppano strutture, tessuti e organi differenti.
[44] Philip Ball. What on earth is a xenobot? AEON, 30 Aug 2022
[45] Ibidem
[46] Ibidem
[47] Ibidem
[48] Gli attrattori possono contenere insiemi invarianti. Un insieme limite è l’insieme degli stati in cui entra un sistema dopo un periodo di tempo infinito.
[49] Douglas Blackiston, Emma Lederer, Sam Kriegman, Simon Garnier, Joshua Bongard and Michael Levin. op. cit. 17 March 2021 / Sam Kriegman, Douglas Blackiston, Michael Levin and Josh Bongard. op. cit. Nov. 29, 2021
[50] Philip Ball. What on earth is a xenobot? AEON, 30 Aug 2022
[51] Ibidem
[52] Ibidem
[53] Genoma: totalità dei cromosomi contenuti in una cellula.
[54] Metazoi: raggruppamento sistematico comprendente tutti gli animali che presentano una condizione monocellulare allo stadio di germe ma che raggiungono durante lo sviluppo una condizione pluricellulare con un numero più o meno elevato di elementi somatici variamente differenziati.
[55] Philip Ball. What on earth is a xenobot? AEON, 30 Aug 2022
[56] Caenorhabditis elegans è un verme nematode fasmidario, lungo circa 1 mm, che vive nel suolo, in regioni temperate. Si tratta di un organismo modello molto usato per lo studio della biologia dello sviluppo e dell’apoptosi. Le prime ricerche su C. elegans sono state avviate nel 1962 da Sydney Brenner.
[57] Philip Ball. What on earth is a xenobot? AEON, 30 Aug 2022
[58] Ruiz-Trillo Iñaki and Nedelcu Aurora M. op. cit. 2016
[59] Ibidem
[60] Dictyostelium discoideum ameba appartenente al genere Dictyostelium, molto studiata come organismo modello in biologia dello sviluppo, biologia molecolare e recentemente in tossicologia ambientale. Come le altre specie appartenenti a questo genere, la d. discoideum è un’ameba unicellulare che vive nello strato di terriccio e foglie in decomposizione del sottobosco, nutrendosi essenzialmente di batteri. Nel caso in cui il cibo inizi a scarseggiare o comunque le condizioni ambientali diventino avverse, una moltitudine di amebe si aggregano rispondendo a stimoli chimici mediati da cAMP. Si origina così una forma pluricellulare talvolta definita pseudoplasmodio, somigliante ad una lumaca, in grado di spostarsi. Lo pseudoplasmodio migra verso zone fortemente illuminate e lì inizia un processo di differenziamento con la formazione una base, uno stelo e un corpo fruttifero nel quale le amebe si differenziano in spore, mentre le amebe che costituiscono lo stelo sono destinate a morire per disseccamento. Le spore vengono liberate e, in presenza di un ambiente favorevole, dal rivestimento protettivo emergono individui ameboidi unicellulari. È un Eucariota, che condivide molti percorsi metabolici con gli organismi superiori. Inoltre possiede caratteristiche peculiari che lo rendono un buon modello per lo studio di: chemiotassi, differenziamento cellulare, trasduzione del segnale e fagocitosi. La forma unicellulare possiede un corredo cromosomico aploide sequenziato interamente dal genetista Ludwig Eichinger. Contiene approssimativamente 12.500 geni su 6 cromosomi (per raffronto, il genoma umano ha 20.000-25.000 geni su 23 cromosomi). Presenta un alto livello dei nucleosidi adenosina e timina (~77%). Il genoma completo è di circa 34 Mb. Fonte: Ludwig Eichinger e Angelika A. Noegel, Crawling in to a new era – the Dictyostelium genome project. in EMBO Journal, vol. 22, n. 9, pp. 1941-1946, 2003
[61] Ruiz-Trillo Iñaki and Nedelcu Aurora M. op. cit. 2016
[62] C. owczarzaki è un eucariota unicellulare che occupa una posizione filogenetica chiave nella nostra comprensione dell’origine della multicellularità animale, come uno dei parenti unicellulari più vicini agli animali. Fonte: Suga, Hiroshi; Chen, Zehua; Mendoza, Alex de; Sebé-Pedrós, Arnau; Brown, Matthew W.; Kramer, Eric; Carr, Martin; Kerner, Pierre; Vervoort, Michel. “The Capsaspora genome reveals a complex unicellular prehistory of animals”. Nature Communications. 4: 2325, 14 Aug 2013
[63] Un fattore di trascrizione è una proteina che lega il DNA in una regione specifica di un promotore o di un enhancer, da dove poi regola la trascrizione. I fattori di trascrizione possono essere attivati o disattivati selettivamente da altre proteine, spesso come passo finale della trasduzione.
[64] Ruiz-Trillo Iñaki and Nedelcu Aurora M. op. cit. 2016
[65] Ibidem
[66] Ibidem
[67] In riferimento alla sistematica biologica per quanto riguarda dei caratteri fenotipici primitivi, già presente nell’antenato comune di linee filetiche diverse.
[68] Ruiz-Trillo Iñaki and Nedelcu Aurora M. op. cit. 2016
[69] Philip Ball. What on earth is a xenobot? AEON, 30 Aug 2022
[70] Ibidem
[71] Michel Morange. A History of Biology. Princeton University Press, 2021
[72] Craig B. Lowe, Manolis Kellis, Adam Siepel, Brian J. Raney, Michele Clamp, Sofie R. Salama, David M. Kingsley, Kerstin Lindblad-Toh, and David Haussler. Three Periods of Regulatory Innovation During Vertebrate Evolution. Science, Vol. 333, Issue 6045, pp. 1019-1024, 19 Agu 2011
[73] Ibidem
[74] Gli esoni costituiscono, insieme agli introni, la porzione di un gene (eucariotico o di archeobatteri) che viene trascritta dalle RNA polimerasi durante il processo di trascrizione. Gli esoni, in seguito al processo di splicing del trascritto primario, detto hnRNA, si ritrovano negli mRNA maturi; talvolta il trascritto primario può subire un processo di splicing alternativo in seguito al quale, negli mRNA maturi, si possono ritrovare anche gli introni o parti di essi. Spesso, erroneamente, viene affermato che gli esoni costituiscano la parte codificante del genoma; questo sarebbe vero solo in parte: se, infatti, risulta vero che tutta la sequenza che codifica una proteina deve risiedere su uno o più esoni, non sarebbe invece sempre vero che un esone sia codificante: esoni non codificanti sono stati individuati in molti geni umani. Nel DNA degli Eucarioti un gene è composto da un certo numero di esoni e di introni; questi ultimi possono essere di dimensioni molto varie anche dell’ordine di centinaia di migliaia di basi. Molte fasi dei meccanismi di rimozione degli esoni durante la maturazione dell’mRNA sarebbero ancora sconosciute mentre alcuni passaggi comincerebbero ad essere noti: ad esempio generalmente gli esoni sono delimitati da due coppie di nucleotidi, un GT e un AG, ma questo da solo non sarebbe sufficiente a definire l’esone. Si tratterebbe comunque di un processo estremamente preciso, che dovrebbe identificare un piccolo esone di qualche decina o centinaia di basi in mezzo a una regione cromosomica che può essere di centinaia di migliaia di basi e inoltre se questi meccanismi vengono meno, come succede ad esempio quando le sequenze GT e AG vanno incontro a mutazione, gli esoni non verrebbero riconosciuti in maniera appropriata e ne deriverebbe la produzione di un RNA anomalo, che non sarebbe in grado di produrre la proteina normale: sarebbe quanto succede in numerosi pazienti affetti da malattie genetiche. L’mRNA maturo, negli Eucarioti, sarebbe di solito costituito oltre che dalla parte codificante (Coding DNA sequence o CDS), anche da due regioni non tradotte (UTR) poste una al 5′ e l’altra al 3′ del trascritto. Questo comporterebbe quindi l’esistenza di esoni completamente o parzialmente non codificanti, ossia quelli che andranno a costituire le UTR del trascritto maturo. Fonte: James D. Watson, Biologia molecolare del gene, 6ª ed., Zanichelli, pp. 416-417, 2009
[75] Craig B. Lowe, Manolis Kellis, Adam Siepel, Brian J. Raney, Michele Clamp, Sofie R. Salama, David M. Kingsley, Kerstin Lindblad-Toh, and David Haussler. op. cit. 2011
[76] Ibidem
[77] Ibidem
[78] Philip Ball. What on earth is a xenobot? AEON, 30 Aug 2022
[79] Craig B. Lowe, Manolis Kellis, Adam Siepel, Brian J. Raney, Michele Clamp, Sofie R. Salama, David M. Kingsley, Kerstin Lindblad-Toh, and David Haussler. op. cit. 2011
[80] Philip Ball. What on earth is a xenobot? AEON, 30 Aug 2022
[81] Craig B. Lowe, Manolis Kellis, Adam Siepel, Brian J. Raney, Michele Clamp, Sofie R. Salama, David M. Kingsley, Kerstin Lindblad-Toh, and David Haussler. op. cit. 2011
[82] Ibidem
[83] Ibidem
[84] Il Cambriano: una delle più importanti divisioni della scala dei tempi geologici che comincia 541,0 ± 1,0 milioni di anni fa, alla fine dell’eone Proterozoico, e si conclude 485,4 ± 1,9 milioni di anni fa con l’avvento del periodo Ordoviciano. Primo periodo dell’era paleozoica nell’eone Fanerozoico. Fonte: Amthor, J. E. et al., Extinction of Cloudina and Namacalathus at the Precambrian-Cambrian boundary in Oman. Geology, 31: pp 431–434, 2003
[85] Philip Ball. What on earth is a xenobot? AEON, 30 Aug 2022
[86] Sam Kriegman, Douglas Blackiston, Michael Levin and Josh Bongard. op. cit. Nov. 29, 2021
[87] Ibidem
[88] La palatoschisi è una fessura o spaccatura nel palato (o tetto della bocca) presente sin dalla nascita. Durante lo sviluppo embrionale, all’interno del grembo materno, il tessuto che forma il palato non si unisce completamente, lasciando un’apertura, detta schisi, sinonimo di interruzione.
Articoli correlati:
 L’apprendimento automatico metterebbe in evidenza l’infatuazione dell’Occidente per la certezza come segno di conoscenza
L’apprendimento automatico metterebbe in evidenza l’infatuazione dell’Occidente per la certezza come segno di conoscenza
 Decostruzione del termine Intelligenza Artificiale.
Decostruzione del termine Intelligenza Artificiale.
 L’obsolescenza della metafora del cervello come computer
L’obsolescenza della metafora del cervello come computer
 Bruno Latour: La ridefinizione delle scienze.
Bruno Latour: La ridefinizione delle scienze.
 Di animali e umani: diventare centauro
Di animali e umani: diventare centauro