


BIO – Medicina Costruzione Sociale nella Post-Modernità – Educational Papers • Anno V • Numero 19 • Settembre 2016
Scritto in collaborazione con Eugenia D’Alterio – biologa
Preliminari
Il nostro mondo soggettivo (e, certamente, anche quello prevedibilmente oggettivo) è pieno delle varietà della vita. Immaginiamo, ad esempio, di fare una passeggiata attraverso un parco con viali alberati. Alberi che gettano piacevoli ombre oscillanti sotto il sole di mezzogiorno. Noteremmo uccelli e farfalle librare nell’aria e, forse, uno scoiattolo arrampicarsi agilmente su un tronco d’albero e, inoltre, una miriade di insetti districarsi per terra o nel fogliame o volando nell’aria.
Prendiamoci un momento per esaminare ciò che ci circonda come con l’occhio di un esperto biologo. Al suo meglio, la scienza ci rivela i tanti livelli e modelli di realtà che si nascondono dietro la nostra esperienza “macroscopica” e giornaliera del mondo. Seguendo il paradigma scientifico di interpretazione della realtà, la scena del parco ci porterebbe a realizzare qualcosa di veramente notevole. Esaminando piante e animali vari che ci circondano, è stato documentato che noi umani siamo direttamente correlati ad ogni altro essere vivente sulla Terra. Ma, con un po’ di osservazione critica, potremmo scoprire questa relazione da soli, una relazione che non è né metafisica né teologica ma, prettamente, di natura chimica-biologica.
Un’osservazione parziale è sufficiente a dimostrare che se siamo, biologicamente, strettamente correlati ai nostri genitori e ai nostri fratelli e sorelle e famiglia tutta, lo siamo, anche, agli altri uomini e donne che potrebbero trovarsi lungo il nostro viale alberato. Certamente, siamo tutti fisicamente trascurabilmente diversi, variando solo in sfumature di proporzione e sottigliezze d’ombra, condividiamo gli stessi sensi, camminiamo e parliamo con la stessa combinazione di muscoli e ossa, siamo tutti nati nello stesso modo (o quasi) e moriremo tutti nella misura in cui i nostri corpi si consumano dopo un periodo di tempo simile (incidentalità, disabilità e malattie a parte). E non è neanche necessario tracciare alberi genealogici per dimostrare che tutti gli esseri umani sono legati: ciò si può affermare solo guardandoci, non per niente apparteniamo alla stessa specie: quella umana.
Il rapporto con il nostro più vicino parente biologico di altre specie, tuttavia, richiede un po’ più di osservazione per essere identificato. Un viaggio allo zoo rivelerà la nostra stretta parentela con animali molto familiari come gli scimpanzé o gorilla. Ma uccelli e mammiferi minori, rettili, anfibi, pesci e microrganismi vari sono stati tutti “tappe” biologiche di una “parentela” remota. Un breve studio della fisiologia è necessario per dimostrarne la somiglianza ancestrale. Tutti noi condividiamo apparati e organi, quali un sistema digestivo e nervoso, una struttura ossea e muscolare e quant’altro costituisce la nostra fisicità. Le differenze tra noi e gli elefanti o le lucertole sono inezie di grandezza e riguardano specializzazioni in un processo evolutivo (adattivo) che ha visto coinvolti tutti i viventi.
La questione della somiglianza però diventa davvero sempre più interessante quando si inizia a guardare la nostra “famiglia” allargata dal punto di vista micro-biologico. Questa include tutti gli esseri viventi: ogni sorta di animale o pianta o microrganismo, dal più grande al più piccolo. Certo, è necessario utilizzare gli strumenti della biologia per vedere questa nostra relazione con tutti i viventi. Uno studio di fisiologia ci evidenza, ad esempio, le differenze di specie, ma quando si guarda al microscopio, troviamo che tutti gli organismi viventi sono costituiti da cellule e che tutte le cellule appaiono sorprendentemente simili. La cellula è l’ unità morfo-funzionale di tutti gli organismi viventi che con i suoi componenti, molecole in primis, orchestra la vita di uni e pluricellulari.
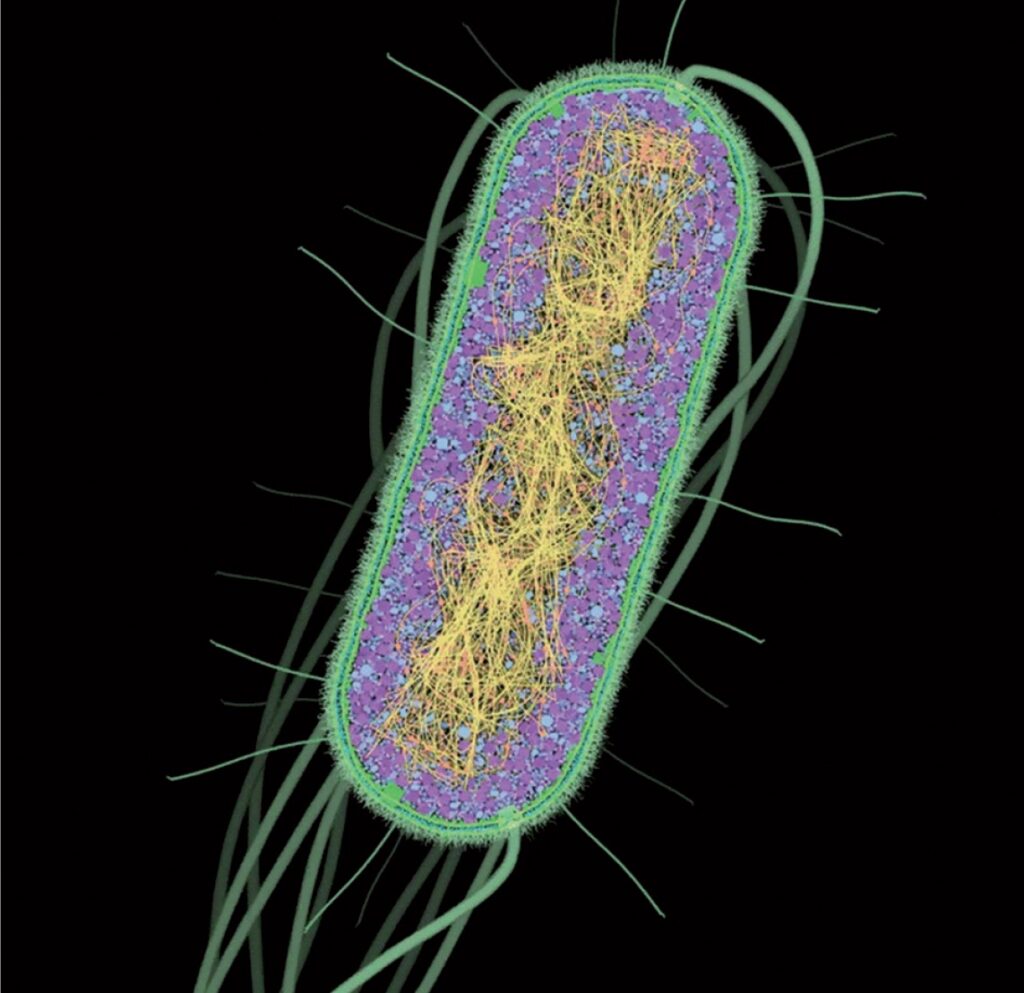
Seguendo il paradigma scientifico tutto ciò che chiamiamo biologicamente vita sulla Terra è composto da cellule che sono, a loro volta, composte da molecole. L’immagine mostra una sezione trasversale di una cellula batterica. Essa è circondata da una parete cellulare multistrato di colore verde. I lunghi flagelli a forma di cavatappi sono attivati da “motori” nella parete cellulare, spingendo così la cellula attraverso il suo ambiente. L’interno della cellula è pieno di “sistemi” molecolari per la costruzione e la riparazione di molecole, per sfruttare diverse fonti di energia e per il rilevamento e la protezione contro i pericoli ambientali (70.000 X). Fonte: Goodsell David S. The Machinery of Life. Springer, New York, 2010, p. 1
Forse la più notevole delle osservazioni della biologia è che soprattutto i microrganismi (batteri in particolare) condividono questa storia di familiarità (origine) ancestrale. I batteri sono costituiti da un’unica cellula (Immagine 1.1) al posto delle migliaia di miliardi di cellule che compongono il nostro corpo, ma quella singola cellula ha, in linea di massima, gli stessi meccanismi delle nostre cellule specializzate. Infatti, se si guardano molto da vicino le molecole cellulari che orchestrano il processo della vita, la somiglianza è evidente (immagine 1.2). Ogni essere vivente sulla Terra utilizza un insieme simile di molecole per nutrirsi, respirare, muoversi e riprodursi1. A causa di questo, qualsiasi essere vivente uni o pluricellulare che sia richiede acqua e nutrimento e, potenzialmente, soccombe in condizioni avverse.
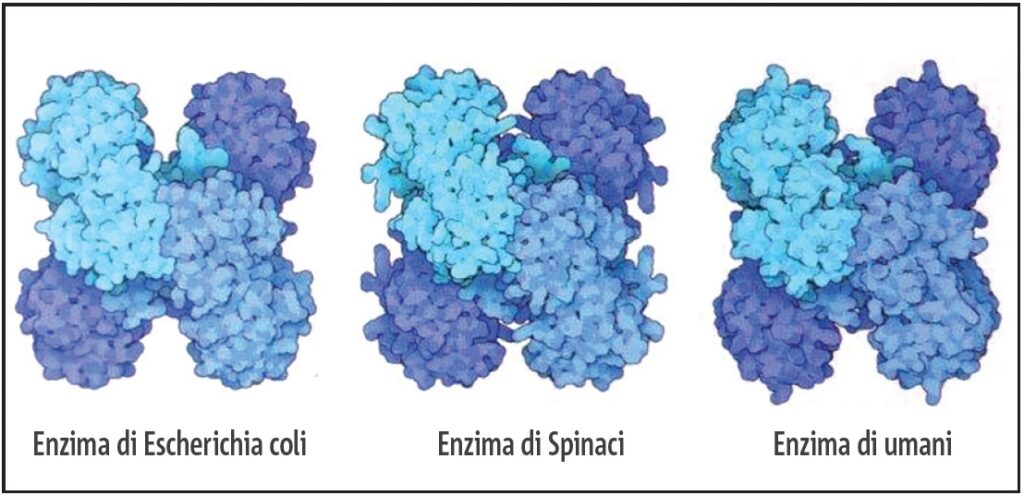
Molti meccanismi molecolari sono praticamente identici in tutte le cellule viventi. Ciò è particolarmente vero per le molecole che svolgono un ruolo fondamentale nel processo della vita, come l’enzima gliceraldeide-3-fosfato deidrogenasi che è essenziale per il metabolismo degli zuccheri in tutti e tre gli organismi. Questa figura mostra la forma simile dell’enzima da cellula batterica (a sinistra), cellula vegetale (al centro), e cellule umane (a destra) (5,000,000X) Fonte: Goodsell David. S. The Machinery of Life. Springer, New York, 2010, p. 3
In questo articolo andremo ad esplorare questo “processo di nascita comune” dei meccanismi molecolari. Inizieremo con uno sguardo agli stessi e all’insolito mondo molecolare – cellulare in cui operano. Si farà un breve accenno alle dimensioni molecolari e cellulari per comprendere meglio gli ordini di grandezza che andiamo ad esaminare.
Una questione di scale
Quasi tutto ciò di cui parleremo in questo articolo è troppo piccolo per essere visto a occhio nudo, poiché al di sotto del nostro potenziale visivo, ma immaginiamo di poterlo fare e pensare al tipo di realtà che potremmo osservare: anticorpi che attaccano un virus, segnali elettrici scendere giù per le fibre nervose, proteine che costruiscono nuovi filamenti di DNA e infinite altre realtà. Se così fosse, molte delle domande rivolte agli attuali standard della scienza sarebbero risposte a colpo d’occhio. Ma il mondo delle molecole, un mondo a scala nano-metrica2, è separato dai nostri modelli percettivi quotidiani dell’esperienza da una scoraggiante differenza di milioni di volte inferiori in termini di dimensioni, infatti il mondo delle molecole è completamente “invisibile” alla nostra percezione naturale3. Di conseguenza, la maggior parte di noi, se non specialisti del settore, ha un concetto totalmente soggettivo della dimensione relativa ad una visione del mondo infinitesimale come quella attinente, ad esempio, alle molecole d’acqua, alle proteine, ai ribosomi, ai batteri, per citarne alcune.
Le cellule sono piccole, ma non così piccole come le molecole che esse contengono. Le dimensioni delle cellule sono variabili e dipendono dai fattori funzionali (organismi unicellulari o pluricellulari con cellule specializzate) e sono, in genere, comprese nell’ordine dei micrometri (milionesimo di metro e millesimo di millimetro, simbolo: μm, 1μm = 10-6m), tra 10 μm e 50 μm4. Questo ordine di grandezza ci dà un’idea di quanto, in termini di dimensioni, sia realmente non percepibile dal nostro campo visivo (escludendo, ovviamente, le uova di uccelli e rettili) nel nostro mondo quotidiano. Cellule più grandi, come quelle dei protozoi, possono essere viste con una lente di ingrandimento (bassa risoluzione) ma è necessario un microscopio ad alta risoluzione per esaminare le altre. Le cellule degli organismi unicellulari presentano caratteri morfo-funzionali solitamente uniformi ma con l’aumentare del numero di cellule di un organismo, come quello umano, invece, le cellule che lo compongono si differenziano e si specializzano in forma, grandezza, rapporti e funzioni fino alla costituzione di tessuti ed organi. Le dimensioni delle cellule umane variano dalle più piccole di 5 – 7 μm (eritrociti e linfociti) a quelle medie di 10 – 20 μm (quasi tutte) e quelle più grandi di 80 – 200 μm (neuroni, adipociti, ovociti). Il numero di cellule che compone, grosso modo, il corpo umano è di 37.200 miliardi, secondo recenti studi. Prendiamo, ad esempio, un chicco di riso, nelle varie classi, mediamente lungo tra i 5 e 6 mm, quindi 5000- 6000 volte più grande rispetto al micron e immaginiamo una nostra stanza piena di questi chicchi. Ce ne vorrà una quantità smisurata e ciò ci dà un’idea dei miliardi di cellule che compongono una minima parte del nostro corpo5.
Al di sotto delle dimensioni cellulari c’è il mondo delle molecole. Le molecole sono così piccole che sono più piccole della lunghezza d’onda della luce, quindi non c’è modo di “vederle” con la risoluzione di un microscopio ottico. La microscopia è per definizione una tecnica che consente di visualizzare oggetti così piccoli da essere fuori portata per l’occhio umano. Per molte applicazioni, il campione può essere illuminato e visualizzato sfruttando la porzione ottica dello spettro elettromagnetico ma quando ci si spinge a dimensioni ancora inferiori s’incontra un limite fondamentale, dovuto alla natura ondulatoria della luce: non è possibile distinguere due oggetti che sono separati da una distanza inferiore alla metà della lunghezza d’onda della luce incidente. Lo spettro ottico è compreso indicativamente tra i 400 e i 700 nanometri (miliardesimi di metro), corrispondenti, rispettivamente, al limite dell’ultravioletto e dell’infrarosso. Poiché l’ordine di grandezza delle dimensioni di una molecola possono variare da decine di nanometri a migliaia di nanometri, molte molecole più piccole o i particolari di quelle più grandi sono oltre le possibilità della microscopia ottica. Per questa ragione usiamo metodi come la cristallografia a raggi X, la spettroscopia NMR, la microscopia elettronica ad effetto tunnel (STM) che sfrutta un effetto quantistico di passaggio della corrente di elettroni tra il campione e una punta di dimensione nanoscopica e il microscopio a forza atomica (AFM) in cui la stessa punta nanoscopica interagisce con il campione tramite le forze di Van der Walls, passando sul campione come una puntina su un vecchio disco in vinile6, tutto questo per scoprire la disposizione degli atomi nelle molecole per, poi, creare “modelli visivi” (immagine 1.3). Ricordiamo che l’ordine di grandezza delle molecole è in nanometri (miliardesimo di metro o milionesimo di millimetro: simbolo nm ovvero, 1 × 10-9 m), mentre per le cellule è in micrometri (milionesimo di metro e millesimo di millimetro: simbolo μm, ovvero, 1 × 10-6 m), per cui un micrometro equivale a 1. 000 nanometri (nm). Da qui il passo è breve per raffrontare dimensioni molecolari a quelle di una tipica cellula.
Due tipi di immagini sono utilizzate per la maggior parte delle illustrazioni in questo articolo. Per creare immagini di singole molecole, come la molecola di emoglobina mostrata, un programma per computer disegna una sfera per ogni atomo nella molecola, centrata sul nucleo dell’atomo e circa le dimensioni della nube di elettroni che circonda il nucleo. In queste illustrazioni si possono facilmente vedere i singoli atomi – questa è disegnata a 5.000.000 X di ingrandimento. Nelle illustrazioni disegnate a mano che mostrano molecole all’interno delle cellule, come la parte del globulo rosso mostrato a sinistra, le forme di queste molecole sono semplificate e i singoli atomi sono troppo piccoli per essere visti (sarebbero circa il formato di un grano di sale in questo ingrandimento). Le illustrazioni disegnate a mano sono presentate in un’amplificazione coerente di 1.000.000 X Fonte: Goodsell David S. The Machinery of Life. Springer, New York, 2010, p. 4
Il mondo molecolare
La molecola è un’entità elettricamente neutra composta da due o più atomi dello stesso elemento o di elementi diversi uniti fra loro da un legame chimico covalente. Viene spesso definita come la più piccola parte in cui può essere suddivisa una sostanza (proteina, ad esempio) conservandone la composizione e le proprietà chimiche che la caratterizzano.
Le molecole nelle nostre cellule svolgono le loro funzioni in un mondo, a noi, quotidianamente sconosciuto e dobbiamo tenerne conto quando ne parliamo. Quando si cerca di capire un processo molecolare, la nostra intuizione può portarci fuori strada poiché i principi che guidano gli oggetti nel nostro mondo di tutti i giorni – gravità, attrito, temperatura – sono differenti su scala molecolare e spesso hanno effetti sorprendentemente diversi.
Di base rimane l’oggettività della solidità della materia. Su scala molecole non abbiamo bisogno di preoccuparci troppo del formalismo della meccanica quantistica che attraverso la caratterizzazione probabilistica di una particella fornita dalla funzione d’onda permette di spiegare la natura elettromagnetica dei legami fisici e chimici che governano il comportamento delle molecole e dei loro costituenti. In prima approssimazione, e per quanto gli studiosi sono riusciti a documentare, possiamo considerare che le molecole hanno dimensione e forme definite ed è, anche, perfettamente ragionevole immaginarle urtarsi e combaciarsi (aderire con legami chimici) se le loro forme funzionali corrispondono. In definitiva, possiamo pensare a loro come oggetti fisici , come tavoli e sedie e, per di più, così piccole che la gravità è del tutto trascurabile. Movimenti e interazioni delle molecole biologiche sono completamente dominati dalle molecole d’acqua circostanti. A temperatura ambiente, una proteina di medie dimensioni viaggia ad una velocità di circa 5 nanometri al secondo. All’interno della cellula, tuttavia, questa proteina circondata da molecole di acqua, rimbalza avanti e indietro, muovendosi in modo casuale da un posto all’altro ma sempre a grande velocità e richiedendo energia per gli spostamenti (immagine 1.4)7.
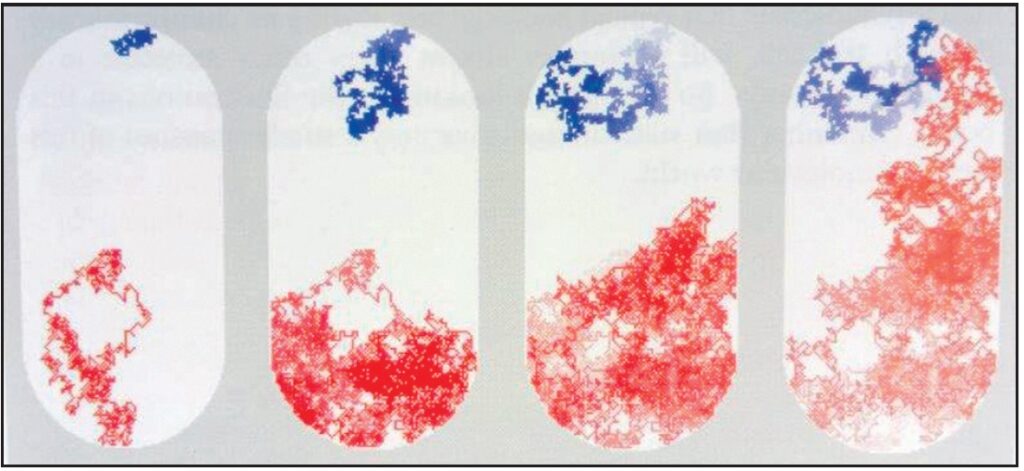
Le molecole costantemente si diffondono nell’interno delle cellule, in modo casuale da un posto all’altro. Quest’illustrazione mostra diverse istantanee da una simulazione al computer di una molecola di una proteina e di una molecola di zucchero che si diffondono all’interno di una cellula batterica. Il percorso della proteina è mostrato in blu e il percorso dello zucchero è mostrato in rosso. Essi partono da estremità opposte e esplorano gran parte dello spazio interno prima di raggiungere l’un l’altro. Fonte: Goodsell David. S. The Machinery of Life. Springer, New York, 2010, p. 5
Immaginiamoci una situazione simile nella dimensione in cui viviamo il nostro mondo quotidiano. Si entra, ad esempio, in un terminale aereo e si vuole raggiungere la biglietteria di un’aerolinea sul lato opposto della sala. La distanza è di alcuni metri. Se la sala è vuota, ci si arriva in una manciata di secondi. Ma immaginiamo, invece, che la stanza sia affollata, piena di persone che cercano di raggiungere disordinatamente i loro rispettivi finestrini. In questo margine di tempo, si può essere spinti e, forse, ritrovarsi al punto di partenza. Questo è simile al percorso, anche se casuale, a cui le molecole attendono nella cellula8.
Potremmo chiederci come, in questo mondo molecolare così caotico, la realtà che percepiamo si sia mai conformata. È vero che il moto è casuale ma è vero, anche, che il movimento è molto veloce rispetto a quello nel nostro mondo visibile. Così, nonostante il movimento a livello molecolare sia casuale e diffusivo, esso è abbastanza veloce per la maggior parte delle funzioni cellulari. Ogni molecola urta, semplicemente, intorno, finché non trova il posto “fisiologicamente giusto”9 ossia idoneo a espletare le sue funzioni.
Per avere un’idea di quanto veloce è questo movimento, immaginiamo una cellula batterica tipica come quella nell’immagine 1.1 e piazziamo un enzima ad una estremità e una molecola di zucchero nell’altra. Essi si imbatteranno in giro e vagheranno per tutta la cellula, incontrando molte molecole lungo la strada. In media, però, ci vorrà circa una frazione di tempo infinitesimale perché quelle molecole si urtino almeno una volta. Questo è veramente notevole: ciò significa che qualsiasi molecola, ad esempio, in una tipica cellula batterica, durante il suo viaggio caotico attraverso la cellula, incontrerà quasi ogni altra molecola in tempi infinitesimali10. Quindi, quando guardiamo l’illustrazione della cellula batterica qui postata, dobbiamo ricordarci che l’immagine statica dà soltanto una singola istantanea di questo mondo molecolare affollato e movimentato. Ma, ricordiamoci che tutti i modelli di simulazione sono sempre solo un’istantanea della complessità del reale in cui viviamo.
Meccanismi molecolari
Il corpo umano è un esempio vivente di come funziona la nanotecnologia. Quasi tutto avviene a livello atomico con la condivisione di elettroni11. Singoli fotoni di luce vengono catturati e utilizzati per dirigere il movimento dei singoli elettroni attraverso circuiti elettrici. Le molecole sono imballate e trasportate, con abilità, su distanze di pochi nanometri. Piccolissimi ingranaggi molecolari, come quello nell’immagine 2.1, orchestrano tutti questi processi della vita su scala nano-metrica. Come macchine evolute dai disegni umani del nostro mondo macroscopico, questi meccanismi molecolari si sono evoluti per assolvere funzioni specifiche in modo efficiente e funzionale. Meccanismi evolutivamente perfezionati al livello degli atomi12.
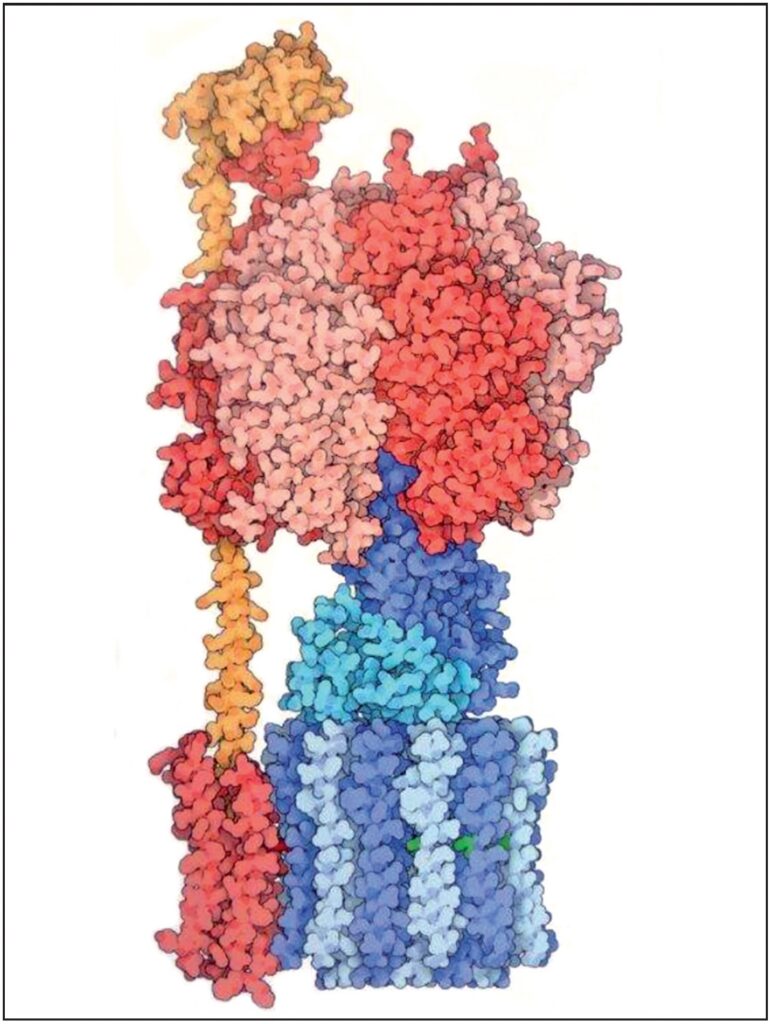
L’ATP sintasi* è un complesso enzimatico o ingranaggio molecolare utilizzato nella produzione di energia chimica. È costruito da oltre 40.000 atomi, ciascuno in un luogo specifico esercitando una funzione specifica (8.000.000 X) Fonte: Goodsell David S. The Machinery of Life. Springer, New York, 2010, p. 8
[ATP sintasi]*13
Le insolite forme organiche dei meccanismi molecolari possono essere viste come scoraggianti e incomprensibili, ma possono essere intese in modo simile, ossia che si sono evolute in modalità adeguate per svolgere una determinata funzione. I meccanismi molecolari naturali, tuttavia, hanno alcune differenze fondamentali riguardo agli artefatti umani di laboratorio ed è necessario acquisire una conoscenza di base di queste differenze per apprezzare, a pieno, ciò che realmente accade su scala molecolare naturale.
Una sfida importante è riconoscere che i complessi meccanismi molecolari sono fatti di atomi. Questo può sembrare ovvio ma, in realtà, provoca problemi. Gli atomi intracellulari sono disponibili solo in poche forme e dimensioni e le cellule fanno quasi tutto il loro lavoro con sei tipi di atomi – carbonio, ossigeno, azoto, zolfo, fosforo e idrogeno – con l’aggiunta di altri atomi solo quando è necessario e per compiti speciali. Questi atomi possono essere collegati in modo molto limitato, definiti dalla chimica sottostante gli atomi stessi. I meccanismi molecolari si sono evoluti all’interno di questi limiti significativi14. Molto simile è cercare di costruire congegni con Meccano o blocchi di Lego: si può costruire una grande varietà di oggetti diversi, ma la forma finale è modellata e limitata dalle forme e dalle connessioni rigide delle unità sottostanti. Vedremo, invece, che i complessi molecolari approfittano di ogni modalità per utilizzare al meglio la loro limitata serie di materie prime.
Le cellule utilizzano, fondamentalmente, quattro “piani” di base nella combinazione degli atomi che compongono i meccanismi molecolari. Così, mentre i nostri artefatti quotidiani sono costruiti in materiale inerte – metallo, legno, plastica e ceramica o quant’altro, i meccanismi su scala nanometrica delle cellule sono materiali biologici costituiti, fondamentalmente, da proteine, acidi nucleici, lipidi e polisaccaridi. Ciascuno di questi “piani”, evoluti nella storia ancestrale cellulare, hanno una caratteristica chimica unica adatta ad un ruolo specifico nella cellula. Due concetti di base sono necessari per capire come questo profilo chimico si manifesta: complementarità chimica (riconoscimento altamente specifico come, ad esempio per le basi azotate del doppio filamento del DNA) e le caratteristiche idrofobiche15 o idrofile dei complessi biologici nei legami intermolecolari.
Così, quando le molecole entrano in contatto, possono interagire tra loro16. Nella maggior parte dei casi, l’interazione non è forte, piuttosto le molecole urtano tra di loro e continuano la loro strada. Tuttavia, ad esempio, se l’interazione è complementare, si legano l’una all’altra. Le molecole interagiscono attraverso una serie di specifiche interazioni di legami. La maggior parte sono interazioni singolarmente deboli, ma poi addensandosi diventano importanti quando una gran parte delle molecole si adatta perfettamente (ossia riconosce) ad una porzione di forma simile di una molecola vicina (Immagine 2.2). I meccanismi molecolari, inoltre, usufruiscono di due tipi di interazioni intermolecolari specifiche: legami a idrogeno tra un atomo di idrogeno e un atomo di ossigeno o di azoto e “ponti di sale” tra atomi che trasportano cariche elettriche opposte. Queste interazioni specifiche agiscono come piccoli elementi di fissaggio per bloccare le molecole insieme17.
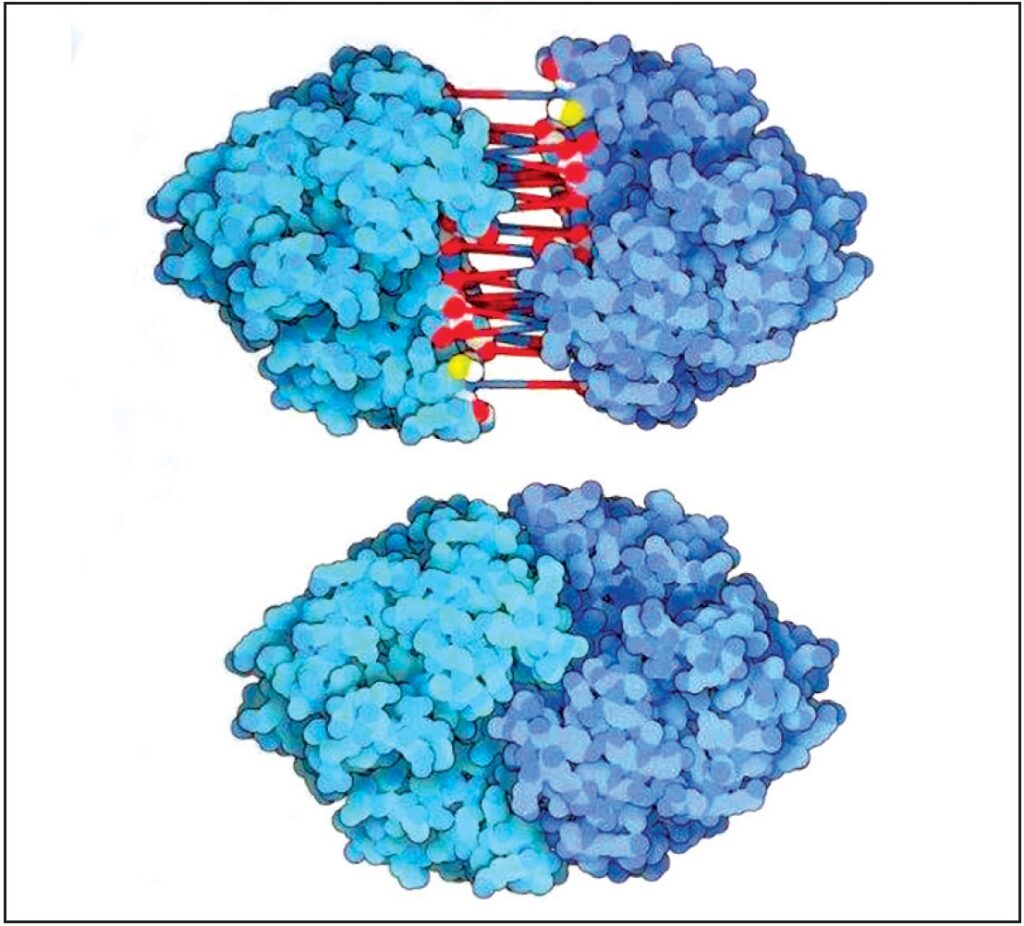
Le molecole biologiche interagiscono attraverso grandi zone complementari sulla loro superficie. L’enzima enolasi, che esegue un passo nel metabolismo degli zuccheri, viene mostrata. Essa è composta da due sub-unità che si combinano per formare gli ingranaggi molecolari attivi. L’immagine mostra le due sub-unità separate, con linee che connettono gli atomi che formano i legami a idrogeno. Si noti come le due forme si connettono e si sposano perfettamente quando si riuniscono (10.000.000 X) Fonte: Goodsell David S. The Machinery of Life. Springer, New York, 2010, p. 11
Il legame a idrogeno o ponte a idrogeno è un caso particolare di forza intermolecolare (dipolo- dipolo) in cui è implicato un atomo di idrogeno coinvolto, a livello molecolare, in un legame covalente con elementi molto elettronegativi (come fluoro (F), ossigeno (O), azoto (N)),[1] i quali attraggono a sé gli elettroni di valenza, acquisendo una parziale carica negativa (δ-) ma lasciando l’idrogeno con una parziale carica positiva (δ+). Contemporaneamente l’idrogeno viene attratto da un atomo elettronegativo di una molecola vicina. Il legame idrogeno presente nelle proteine e negli acidi nucleici è una delle forze che tiene uniti i due filamenti del DNA18.
L’idrofobicità è un concetto più sfuggente che cerca di descrivere la proprietà fisica di specie chimiche (ad esempio molecole) di essere respinte dall’acqua. Si utilizza inoltre il termine in senso più ampio per indicare la proprietà di materiali di non assorbire e non trattenere acqua al loro interno o sulla loro superficie. Infatti, le molecole tendono ad interagire in due modi con l’acqua. Da un lato, le molecole che interagiscono fortemente con essa, spesso molecole ricche di atomi di ossigeno e di azoto, sono chiamate idrofile (ossia che si legano all’acqua). Le molecole idrofile (solubili) sono polarizzate elettricamente e in grado di interagire con l’acqua stabilendo legami idrogeno. Ad esempio, lo zucchero da tavola e l’acido acetico sono piccole molecole idrofile che ci sono familiari. D’altro lato, le molecole neutre e apolari che sono ricche di atomi di carbonio non interagiscono bene con l’acqua e sono chiamate idrofobiche (ossia che non si legano all’acqua). Queste ultime, quando vengono in contatto con l’acqua, tendono a raggrupparsi insieme, raccogliendosi in forme globulari isolandosi dall’acqua circostante (Immagine 2.3). Questo è ciò che accade con l’olio vegetale in acqua: si formano goccioline per minimizzare il contatto delle molecole idrofobe dell’olio con l’acqua19.
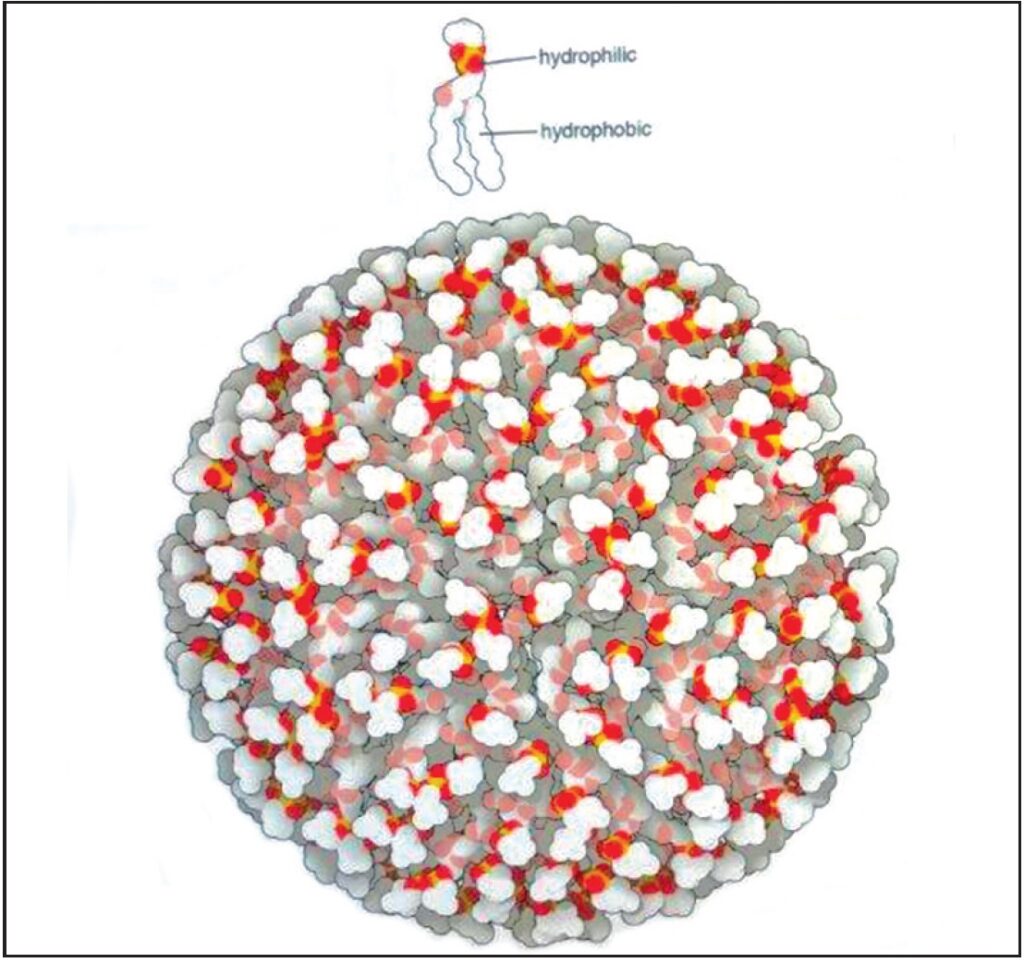
Il fosfolipide mostrato nella immagine ha un gruppo fosfato, mostrato in giallo brillante e rosso, che è idrofilo – e interagisce fortemente con l’acqua, mentre il resto della molecola è composto principalmente da carbonio e idrogeno (colore bianco) ed è idrofobico e interagisce debolmente con l’acqua. Quando i lipidi sono mescolati con acqua, si separano in piccole gocce (o bistrati lipidici, come descritto più avanti, riducendo al minimo il contatto con l’acqua circostante. Nella parte inferiore dell’immagine, molti fosfolipidi si sono fusi in un globulo
compatto, con tutte le parti idrofobe al riparo all’interno. Fonte: Goodsell David S. The Machinery of Life. Springer, New York, 2010, p. 12
I grandi ingranaggi molecolari nelle cellule sfruttano entrambe queste proprietà: idrofilia e idrofobia. Le molecole hanno superfici di forma irregolare che utilizzano legami a idrogeno e i “ponti salini” (l’interazione elettrostatica è responsabile della formazione dei ponti salini tra residui aminoacidici con cariche opposte) per trovare molecole con forme complementari. Spesso le molecole hanno due “regioni”: una idrofila ed una idrofoba che interagiscono, ovviamente, in modo diverso con l’acqua. Diversi modelli di queste “regioni” obbligano le molecole ad agire in modi diversi quando vengono messe in acqua. I quattro “piani” molecolari – proteine, acidi nucleici, lipidi e polisaccaridi – utilizzano diverse combinazioni di queste proprietà per raggiungere diversi obiettivi molecolari e funzionali20.
Acidi nucleici
Gli acidi nucleici (DNA e RNA) sono lunghi polimeri costituiti da un gran numero di nucleotidi legati gli uni agli altri da legami covalenti che contengono, trasportano e decifrano l’informazione genetica. Le molecole di DNA (acido desossiribonucleico) si presentano in doppia elica mentre l’RNA (acido ribonucleico), normalmente, in unico filamento. Svolgono un ruolo essenziale – alcuni direbbero un ruolo centrale – nei processi della vita, in quanto sede dell’informazione genetica, informazione (ereditaria) necessaria per mantenere in vita una cellula e riprodurla sia in un organismo unicellulare che pluricellulare e consentire l’ereditarietà. Così, tutte le informazioni che descrivono come fare le proteine, e quando farle, sono memorizzate nei filamenti di acido nucleico di ogni cellula (nucleata o meno)21.
Le interazioni uniche tra le catene di acidi nucleici le rendono ideali per questo ruolo di “bibliotecarie” della vita cellulare. Nel DNA (acido desossiribonucleico) questi nucleotidi sono disponibili in quattro tipologie: adenina (A), timina (T), citosina (C) e guanina (G). I quattro nucleotidi corrispondono perfettamente in una speciale combinazione nel doppio filamento del DNA: A con T e C con G22.
Questa corrispondenza specifica è la base per la capacità degli acidi nucleici di memorizzare e trasmettere informazioni. Proprio come le stringhe di numeri in un dischetto, le informazioni possono essere memorizzate nella sequenza di nucleotidi lungo un filamento di acido nucleico. Per esempio, la sequenza A-T-G è un codice universale che significa “Start”. Questa informazione può essere letta utilizzando diversi legami specifici di idrogeno: un nuovo filamento23 può essere costruito facendo corrispondere nucleotidi di un filamento sull’altro.
Così la sequenza su un filamento A, T, G, risulterà sul suo filamento complementare T,A,C. La replicazione dei filamenti si ripeterà di generazione in generazione24.
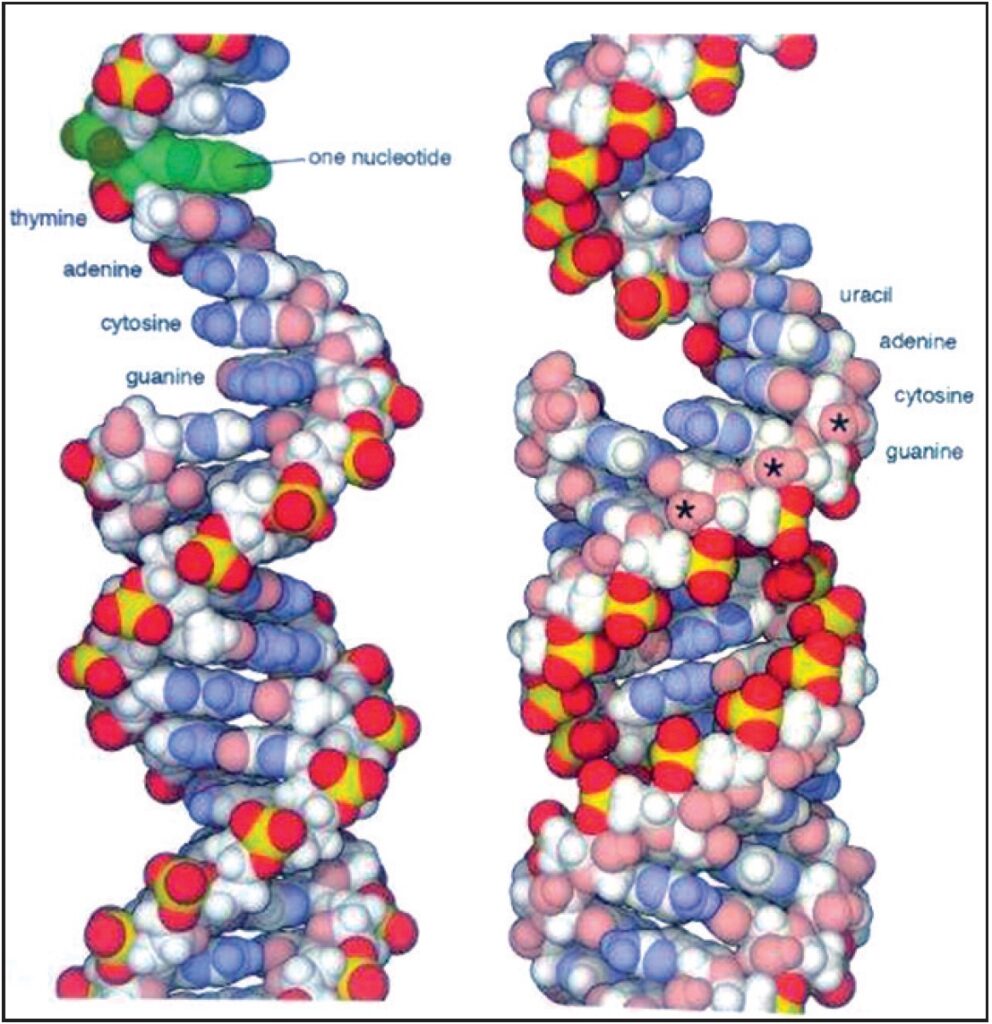
Gli acidi nucleici sono costituiti da lunghe catene di nucleotidi. In questa illustrazione gli atomi di carbonio idrofobi sono di colore bianco, gli atomi idrofili sono leggermente in tonalità pastello (azzurro per l’azoto, rosso per l’ossigeno e giallo per il fosforo). Gli atomi di idrogeno, che sono indicati con sfere più piccole, sono colorati uguali all’atomo a cui sono legati. Il DNA è mostrato sulla sinistra e l’RNA viene visualizzata sulla destra, con doppia elica sul fondo e un singolo filamento in alto. Gli asterischi sul filamento di RNA mostrano alcuni degli atomi di ossigeno presenti nel RNA, ma non nel DNA (20.000.000 X) Fonte: Goodsell David S. The Machinery of Life. Springer, New York, 2010, p. 14
La struttura chimica dei nucleotidi è perfetta per assistere questo trasferimento di informazioni (Immagini 2.4 e 2.5). Ogni nucleotide è composto da una base, che comprende gli atomi di idrogeno tenuti insieme in perfetto orientamento in un anello rigido e un gruppo di zucchero-fosfato che viene utilizzato per collegare i nucleotidi insieme. I gruppi di zucchero-fosfato sono relativamente flessibili, per cui la catena può piegare in molte forme funzionali e i fosfati, altamente caricati, rendono la catena molto solubile in acqua. Le basi, d’altro canto, sono piuttosto idrofobiche e favoriscono forme in cui sono impilate una sopra l’altra, al riparo dall’acqua. Per questo il DNA costituisce la doppia elica: due filamenti si allineano con tutte le basi impilate all’interno e tutti i gruppi fosfato si avvolgono intorno all’esterno25.
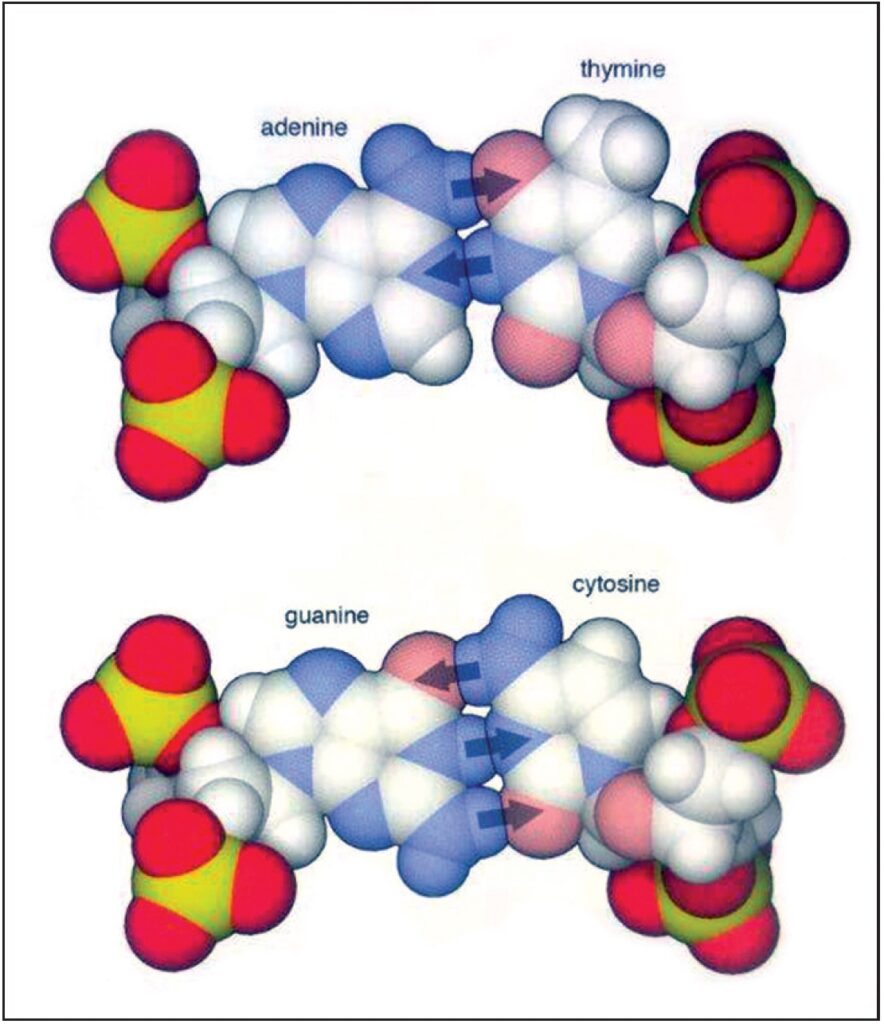
Le basi di DNA interagiscono attraverso una serie di legami idrogeno specifici, combinando adenina con timina e citosina con guanina. L’interazione del legame di idrogeno è più forte quando le basi sono perfettamente allineate side-by-side, combinando un atomo di idrogeno su una base con uno di ossigeno o di azoto sulla base complementare. Adenina e timina forma due dei legami di idrogeno (indicato con le frecce) e guanina e citosina ne formano tre (40.000.000 X) Fonte: Goodsell David S. The Machinery of Life. Springer, New York, 2010, p. 15
Gli acidi nucleici sono costituiti in due varietà nelle cellule: DNA e RNA (immagine 2.6). L’RNA (acido ribonucleico) differisce chimicamente dal DNA in due piccoli modi: lo zucchero ha un atomo di ossigeno supplementare e la base timina è sostituita da uracile, che è più piccolo per un atomo di carbonio e per qualche atomo di idrogeno. Queste piccole differenze chimiche in realtà fanno una grande differenza nella funzione dell’RNA. L’atomo di ossigeno supplementare rende l’RNA un po’ meno stabile del DNA, in modo che il DNA viene utilizzato principalmente come il magazzino centrale di informazioni e l’RNA viene utilizzato per l’elaborazione delle informazioni temporanee. Filamenti di RNA, usa e getta, vengono copiati da modelli di DNA, tagliati, ricoperti, “editati” e trasportati nella cellula per svolgere le loro funzioni, e infine scartati quando il lavoro è fatto26.
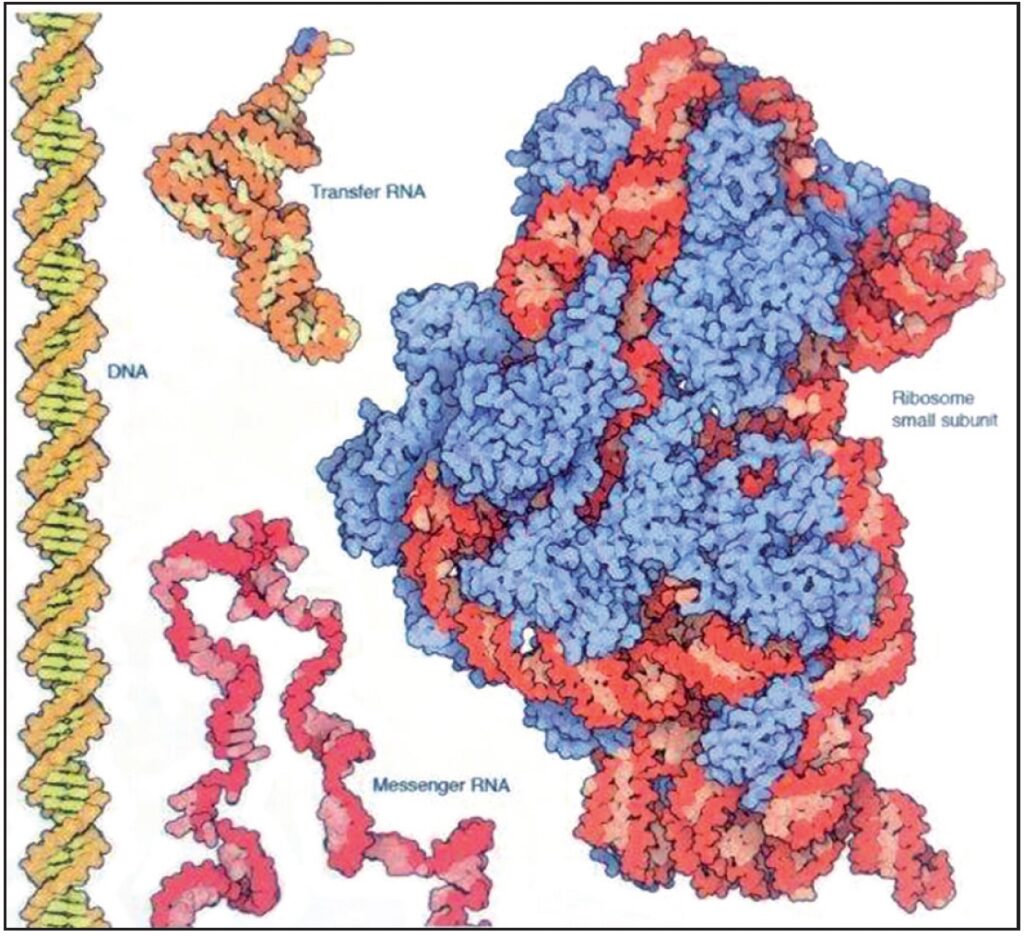
L’acido nucleico esegue diverse operazioni nelle cellule. La doppia elica del DNA è il magazzino principale di informazioni genetiche, e i lunghi filamenti dell’RNA messaggero sono portatori temporanei di queste informazioni. L’RNA di trasferimento e i ribosomi (che sono composti da RNA e proteine) sono ingranaggi essenziali per la sintesi delle proteine (5.000.000 X) Fonte: Goodsell David S. The Machinery of Life. Springer, New York, 2010, p. 16
Gli acidi nucleici, tuttavia, sono troppo limitati nella loro struttura per svolgere i molteplici compiti nella vita quotidiana di una cellula. La chimica delle quattro basi, anche se ideale per il trasferimento di informazioni, sono troppo simili per consentire le diverse reazioni chimiche e meccaniche. Invece, le proteine sono usate per questa loro capacità27.
Proteine
Le proteine o protidi sono macromolecole costituite da lunghe unità di base, gli aminoacidi (20), legati uno all’altro da legami peptidici (ovvero un legame tra il gruppo aminico di un aminoacido e il gruppo carbossilico dell’altro). Le proteine svolgono una vasta serie di funzioni all’interno degli organismi viventi, alcuni esempi sono: una attività enzimatica, ossia agiscono come catalizzatori delle reazioni biologiche , la replicazione del DNA, la risposta agli stimoli e il trasporto di molecole da un luogo ad un altro e molto altro ancora. Le proteine, ovviamente, differiscono l’una dall’altra nella loro sequenza di aminoacidi, che è dettata geneticamente dalla sequenza nucleotidica e che, di solito, si traduce in una struttura proteica tridimensionale specifica che ne determina la sua attività funzionale.
Ovunque in una cellula si vedranno proteine al lavoro, in migliaia di forme e dimensioni specifiche, assemblando in barre, reti, sfere cave e tubi, altre sono motori molecolari che utilizzano energia per ruotare o flettere o strisciare, ma ciascuna con funzione altamente specifica28.
I 20 aminoacidi differenti, di cui sono costituite le proteine, hanno ciascuno una diversa dimensione e carattere chimico (immagine 2.7). Alcuni aminoacidi portano una carica elettrica e interagiscono fortemente con acqua e ioni, altri sono composti principalmente da atomi di carbonio e idrogeno e sono fortemente idrofobici, rendendoli rigidi o molto flessibili. Alcuni possono essere chimicamente reattivi o completamente neutrali. Utilizzando questo diverso “alfabeto” (carattere bio-chimico) dei 20 aminoacidi, le cellule sono in grado di costruire un “vocabolario” ancor più diverso (ampio) di proteine29.
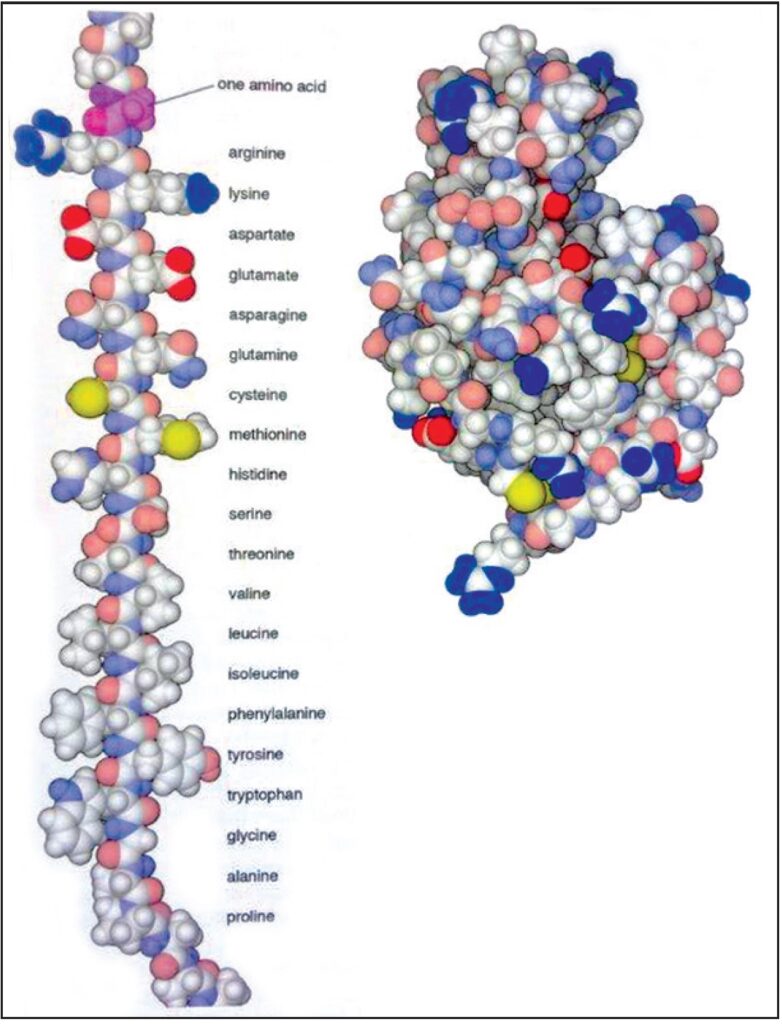
Le proteine sono composte da una catena di aminoacidi che si ripiega in una struttura globulare compatta. Tutti i 20 aminoacidi sono mostrati nella catena estesa su questa fig. 2.7. Si noti la diversa forma e la composizione chimica di ognuna. Le quattro in alto portano una carica elettrica forte, quelle in mezzo sono leggermente cariche e quelle in fondo sono idrofobiche. L’ultima, la prolina, costituisce un nodo rigido nella catena proteica. La piccola proteina lisozima, che contiene 129 amminoacidi, viene mostrata sulla destra (20.000.000 X). Fonte: Goodsell David S. The Machinery of Life. Springer, New York, 2010, p. 18
Quando le catene proteiche sono poste in acqua, esse svolgono un’impresa notevole, effettuando torsioni e pieghe, e trovando una forma ottimale che annidi gli aminoacidi idrofobici all’interno e collochi gli aminoacidi carichi sulla superficie. Sorprendentemente, le proteine sono ingranaggi auto-assemblanti. Questo processo avviene da solo, o con un piccolo aiuto delle proteine “ chaperones” che proteggono la catena pieghevole dall’influenza e intromissione delle proteine confinanti. Così la forma finale della proteina ripiegata è completamente predeterminata dall’ordine degli aminoacidi nella catena proteica30.
Come si può immaginare, solo una piccola frazione delle possibili combinazioni di aminoacidi si piega spontaneamente nella struttura stabile. Infatti, se si assembla una proteina con una sequenza casuale di aminoacidi è probabile che si formerà solo un groviglio casuale e appiccicoso con nessuna funzionalità. Ma le cellule hanno perfezionato le sequenze di aminoacidi nelle loro proteine in tanti anni di selezione evolutiva e gli scienziati stanno scoprendo solo da poco le regole genetiche che guidano il processo di piegatura, che permette di progettare e produrre le (nostre) proteine e renderle funzionali31.
Con il loro ampio potenziale per la diversità, le proteine sono costruite per eseguire la maggior parte delle attività quotidiane della cellula. Un tipico batterio costruisce diverse migliaia di differenti tipi di proteine, ciascuna con una funzione diversa. Le nostre cellule ne costruiscono circa 30.000 tipi diversi, che variano nel formato dai piccoli ormoni proteici, come il glucagone, che ha solo 29 aminoacidi, a enormi proteine come la timina, che ne ha oltre 34.000. La maggior parte delle funzioni cellulari viene espletata da proteine, ovviamente sono necessarie anche tutte le altre molecole biologiche: acidi nucleici, lipidi e carboidrati. Ma le proteine rimangono il jolly di tutti i meccanismi cellulari, pressate a dovere, in innumerevoli forme32.
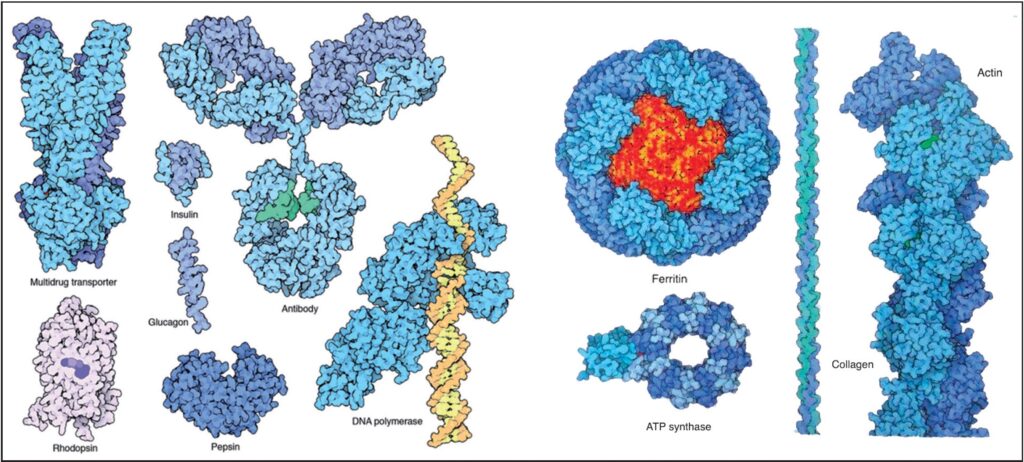
Le proteine si sono sviluppate per molte funzioni diverse. In molti casi, diverse catene proteiche si piegano e, quindi, si associano per formare una struttura più grande.
Seguono alcuni esempi: la proteina di resistenza multi farmaco o P-glicoproteina [MULTIDRUG TRANSPORTER OR P-GLYCOPROTEIN]33 pompa le droghe e i veleni fuori dalle cellule utilizzando un movimento tipo forbice delle sue due catene; la rodopsina è il sensore di luce nelle nostre retine – utilizza una molecola retinale colorata per catturare la luce; l’insulina e glucosio sono ormoni che offrono messaggi opposti sui livelli di zucchero nel sangue; la pepsina è la macchina delle proteine digerenti che rompe il cibo nello stomaco – a causa di questa funzione, deve essere altamente resistente agli acidi; gli anticorpi, specializzati in riconoscimento dei corpi estranei, trovano virus e batteri nel sangue durante le infezioni; l’RNA polimerasi copia le informazioni contenute in un filamento di DNA – l’esempio sposto è da un batterio che vive nelle sorgenti calde, quindi è molto stabile e viene utilizzato nel settore delle biotecnologie per copiare il DNA; la ferritina immagazzina ioni di ferro all’interno delle cellule – è un involucro proteico che circonda una piccola massa cristallina di circa 4000 ioni ferro e di ioni idrossidi. Questa parte della ATP sintasi è un motore rotativo alimentato da energia elettromeccanica. Il collagene forma lunghi cavi robusti che supportano i nostri organi e tessuti, ed è la proteina più comune nel corpo umano; l’actina forma anche travi strutturali, ma quelli che possono essere assemblati e smontati a piacimento (5.000.000 X) Fonte: Goodsell David S. The Machinery of Life. Springer, New York, 2010, p. 20-21
Lipidi
Insieme a glucidi (carboidrati), protidi e acidi nucleici, i lipidi rappresentano una delle quattro principali classi di composti organici di interesse biologico. I lipidi rappresentano un’importante riserva energetica per animali e piante oltre che funzione strutturale (membrane cellulari) e importanti funzioni di regolazione delle attività cellulari (vitamine e ormoni).
I lipidi o grassi sono riconoscibili per la loro proprietà comune di insolubilità. Sono costituiti prevalentemente da atomi di carbonio e di idrogeno uniti tra loro con legami covalenti scarsamente polari (caratteristica che conferisce il comportamento idrofobo) e disposti simmetricamente, solubili in solventi organici come etere dietilico o acetone, alcoli e idrocarburi. Tuttavia, alcuni lipidi presentano, nella loro molecola, gruppi polari (ad esempio i fosfolipidi). I lipidi polari presentano caratteristiche fisico-chimiche peculiari rispetto ai lipidi neutri (apolari). In particolare, i lipidi polari mostrano caratteristiche anfipatiche (solubilità sia in acqua che in solventi apolari) e talora risultano insolubili in solventi organici e solubili in acqua, abilità indispensabile ai vari aspetti funzionali.
Individualmente, i lipidi sono molecole molto piccole, ma quando raggruppate insieme, formano le più grandi strutture della cellula. Poste in acqua, le molecole di lipidi si aggregano per formare foglietti impermeabili. Il doppio foglietto fosfolipidico (spesso denominato bilayer fosfolipidico) è una particolare struttura biologica costituita da fosfolipidi e rappresentata da due strati di fosfolipidi disposti a polarità opposta. Questi foglietti sono usati per racchiudere le cellule, formando il confine primario (la cosiddetta membrana cellulare) che separa l’interno della cellula dall’ambiente esterno e utilizzati anche per costruire compartimenti all’interno delle cellule, come il nucleo e i mitocondri33.
L’interazione insolita dei lipidi con l’acqua li rende così utili. I lipidi, noti comunemente come grassi e oli, sono composti da una piccola “testa” idrofila collegata a due o tre lunghe “code” idrofobe. Quando poste in acqua, le molecole lipidiche spontaneamente si aggregano, imballando le molecole le une affianco alle altre per riparare le lunghe code idrofobe dall’acqua. Questo processo ci è familiare: se si mette una goccia di olio vegetale in acqua, l’olio si aggregherà in gocce nell’acqua (immagine 2.3.).
Poiché i lipidi a due strati (bilayer) sono composti da tante molecole separate, essi sono dinamici e fluidi, permettendo la permeabilità selettiva delle membrane. Dal momento che sono così fluidi, i lipidi a doppio strato rendono le membrane perfette per le cellule. Essendo flessibile la membrana si piega facilmente per soddisfare le esigenze della cellula. Gli eventuali strappi nella membrana sono rapidamente riparati e la membrana può crescere velocemente o ridursi in termini di dimensioni, semplicemente aggiungendo o rimuovendo molecole lipidiche34.
La membrana cellulare è formata in prevalenza da lipidi, in massima parte fosfolipidi, ma nella componente lipidica si vanno a collocare, con importanti funzioni fisiologiche, proteine e una piccola percentuale di glucidi, in forma di glicoproteine e glicolipidi, e di molecole di colesterolo che la stabilizzano. Le proteine di membrana, grazie alla fluidità della componente lipidica, presentano un notevole grado di mobilità; ad esse spetta lo svolgimento della gran parte delle funzioni di membrana separazione dall’ambiente esterno e regolazione del transito di elementi e sostanze chimiche (interno – esterno e viceversa).
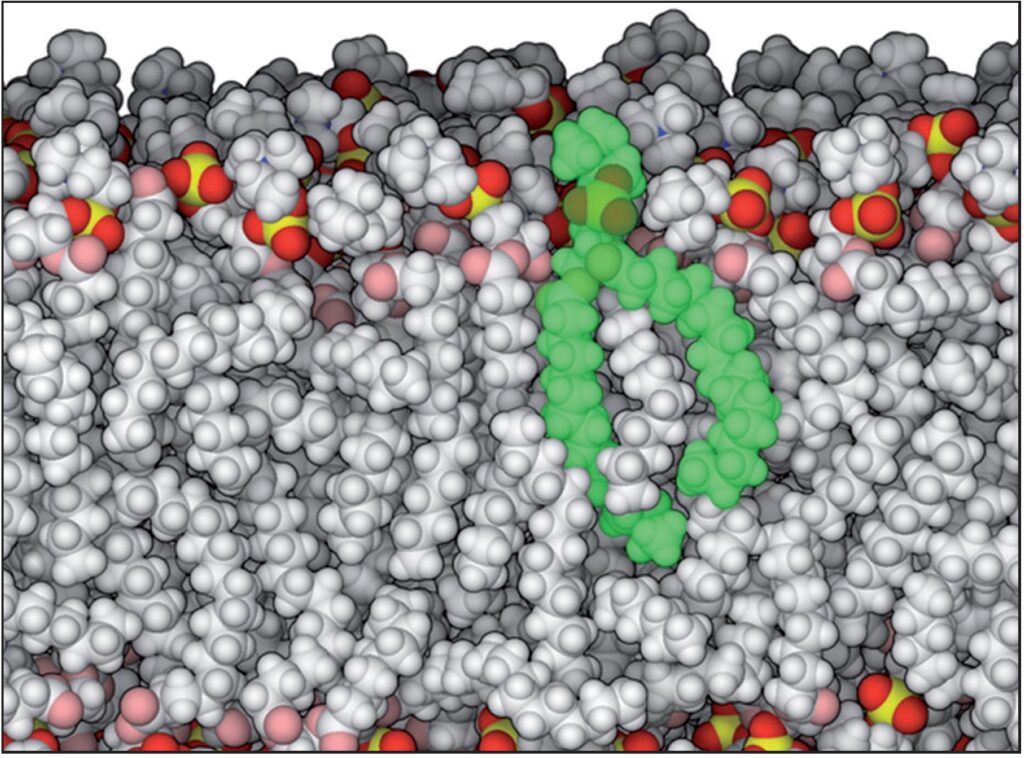
Un lipide (evidenziato in verde) è piccolo rispetto alle proteine e agli acidi nucleici e di gran lunga più idrofobo. Nella cellula, molti lipidi si associano per formare un doppio strato lipidico dinamico. Il doppio strato è illustrato nella sezione esposta rivelando il groviglio di catene di idrocarburi al riparo (20.000.000 X) Fonte: Goodsell David S. The Machinery of Life. Springer, New York, 2010, p. 22
Polisaccaridi
La quarta classe di “materiali” da costruzione di base delle cellule viene utilizzato principalmente per la sua consistenza: i polisaccaridi. Questi sono carboidrati, formati da lunghe catene di monomeri semplici come ad esempio il glucosio. I polisaccaridi di maggior interesse biologico sono rappresentati dal glicogeno, dall’amido e dalla cellulosa. I polisaccaridi si presentano in lunghe catene lineari, spesso ramificate, come le molecole di zucchero tra cui il saccarosio (formato da una molecola di glucosio più una di fruttosio), cioè il comune zucchero da cucina.
Gli zuccheri sono coperti con gruppi idrossilici (un atomo di ossigeno e un atomo di idrogeno) che interagiscono bene con l’acqua e, anche, interagiscono fortemente con altri gruppi idrossilici. I polisaccaridi approfittano di questa proprietà per svolgere due funzioni principali. La prima è di stoccaggio. Lo zucchero, il glucosio in particolare, è la principale fonte di energia utilizzata nella cellula. I polisaccaridi sono usati come il magazzino centrale di questa energia. In tempi di abbondanza, le molecole di zucchero in eccesso sono collegate in grandi granuli di polisaccaridi. In tempi difficili questi granuli sono ripartiti e lo zucchero viene rilasciato. I polisaccaridi sono meno reattivi degli zuccheri individuali e i molti gruppi di idrossile si associano per formare granuli compatti, facilmente immagazzinati, che li rendono migliori per lo stoccaggio di energia rispetto ad un’alta concentrazione di zucchero libero. Nelle piante, il glucosio viene invece immagazzinato sotto forma di amido, lo stesso amido che usiamo per addensare salse e irrigidire i colletti. Negli animali e, quindi, anche nelle nostre cellule, il glucosio è legato, invece, in modo leggermente diverso (polisaccaridi ramificati) per formare glicogeno35.
Se i polisaccaridi rivestono un ruolo importante nelle cellule animali, dove sono utilizzati per costruire alcune delle strutture biologiche più durevoli, da ricordare che molte costruzioni materiali e le pagine di questo trimestrale sono realizzate in gran parte con polisaccaridi, infatti, le fibre di cellulosa da legno, utilizzate allo scopo, sono in gran parte costituite da catene polisaccaridiche lunghe (immagine 2.10). Anche i gusci rigidi degli insetti e delle aragoste, ad esempio, sono fatti di lunghe catene di polisaccaridi: la chitina36.
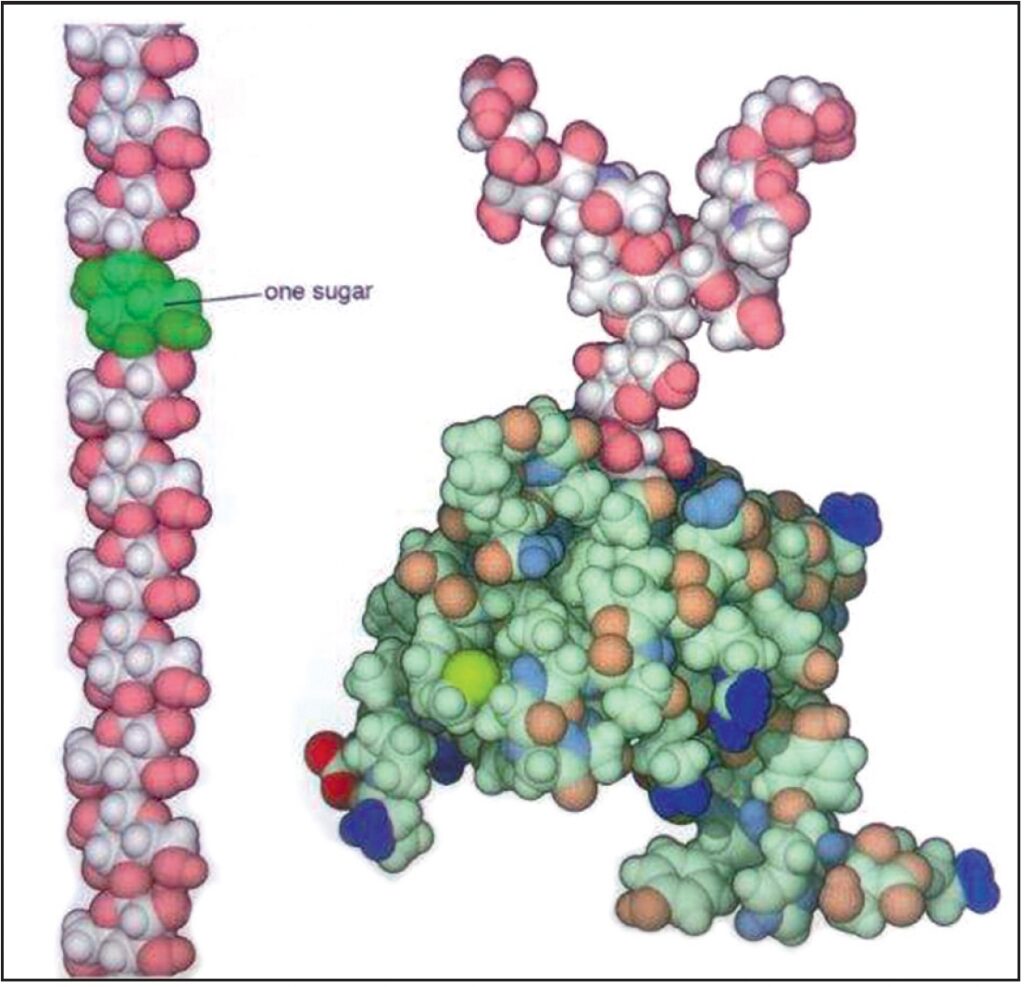
I polisaccaridi sono coperti con gruppi idrossilici (di colore rosa) rendendoli idrofili e solubili in acqua. Due esempi sono mostrati. La cellulosa è mostrata a sinistra, con uno zucchero evidenziato. Un ormone proteico (gonadotropina corionica) è mostrato sulla destra. Esso è composto di una catena proteica corta, tinta in verde, e una catena polisaccaridica ramificata. Piccoli polisaccaridi come questi sono attaccati a molte delle proteine sulla superficie delle nostre cellule (20.000.000 X) Fonte: Goodsell David. S. The Machinery of Life. Springer, New York, 2010, p. 24
Le nostre cellule usano anche polisaccaridi come materiale strutturale, ma molto più piccolo rispetto alla cellulosa o alla chitina. La maggior parte delle cellule sono rivestite con uno strato breve di polisaccaridi legati alle proteine o ai lipidi sulla superficie cellulare. Queste catene si estendono nella cellula e interagiscono con molta acqua. Questa miscela di polisaccaride e acqua forma un “cappotto” colloso intorno alla cellula che funge da barriera protettiva. Per avere un’idea di ciò, come è questa barriera polisaccaride, basta pensare all’ultima volta che abbiamo avuto un raffreddore: il muco deve le sue proprietà distintive alle catene polisaccaridiche attaccate alle sue proteine componenti37.
Il mondo molecolare cellulare
Quello che abbiamo esaminato, sarà riproposto in articoli successivi. Ma è importante aver presentato e illustrato, anche se in maniera molto concisa, che le cellule sono piccoli luoghi affollati, dove molte attività accadono in contemporanea. Abbiamo visto, anche, che i meccanismi molecolari devono svolgere le loro funzioni su scala nanometrica in questo ambiente insolito, ossia nel packaging delle cellule viventi38. Tutto questo comporta una serie di questioni sorprendenti, e qualche nuove opportunità per noi umani.
Le cellule sono incredibilmente “affollate”, tipicamente con 25-35% dello spazio riempito da grandi molecole come le proteine e gli acidi nucleici. Come si può immaginare, queste molecole si imbattono le une contro le altre nel loro spostamento. Questo ha due effetti apparentemente opposti sulla funzione delle molecole. Innanzitutto, le molecole più grandi hanno più difficoltà di spostamento attraverso l’ambiente cellulare, dal momento che sono costantemente bloccate da quelle vicine. Questo rallenta i movimenti molecolari, quindi ci vuole più tempo perché due molecole complementari, ad esempio, si trovino l’una accanto all’altra. Tuttavia, contrastando questo effetto, l’ambiente affollato tende a favorire l’associazione di molecole una volta che esse trovano l’affinità interattiva. Poiché sono costantemente ammassate da molecole confinanti, passano più tempo le une accanto alle altre ed è molto più probabile trovare il corretto orientamento per l’interazione. Questa modalità tende a favorire l’associazione di molecole in grandi complessi, in ambienti affollati, piuttosto che riempire la cellula con un sacco di molecole separate39.
Le cellule sono piene di barriere (reticolo endoplasmatico) che inibiscono o migliorano il movimento delle molecole e sono spesso utilizzate per isolare una sezione della cellula stessa bloccando, quasi completamente, la diffusione delle molecole da una parte all’altra. Questa modalità migliora la funzione delle molecole racchiuse, sequestrandole in un piccolo spazio insieme a tutte le altre molecole necessarie per un particolare compito (funzione). Ma le membrane, di contro, accelerano anche la diffusione delle molecole e l’associazione di alcune di esse. Se una proteina si lega debolmente ad una membrana, essa può legarsi in un punto e poi saltellare sulla superficie della membrana stessa. Poiché la proteina in oggetto avrebbe solo bisogno di muoversi in due dimensioni nello spazio cellulare, su una regione più piccola di spazio la molecola è libera di diffondersi in tre dimensioni. Così, essa può trovare i suoi target (obiettivi) sulla superficie della membrana molto rapidamente. Questo è anche il caso per le proteine che si legano al DNA, infatti, esse possono muoversi rapidamente su e giù le singole eliche trovando la posizione corretta. Per esempio, la proteina repressore lac (che impedisce la trascrizione di alcuni geni) mostra un legame debole e non specifico per la maggior parte delle sezioni di DNA e un legame molto forte vincolante per la particolare sequenza di nucleotidi per cui è progettata a reprimere. All’interno delle cellule, facendo scorrere su e giù un’elica del DNA, la proteina in causa trova i suoi siti di legami preferiti centinaia di volte più velocemente del previsto con una diffusione casuale e tridimensionale40.

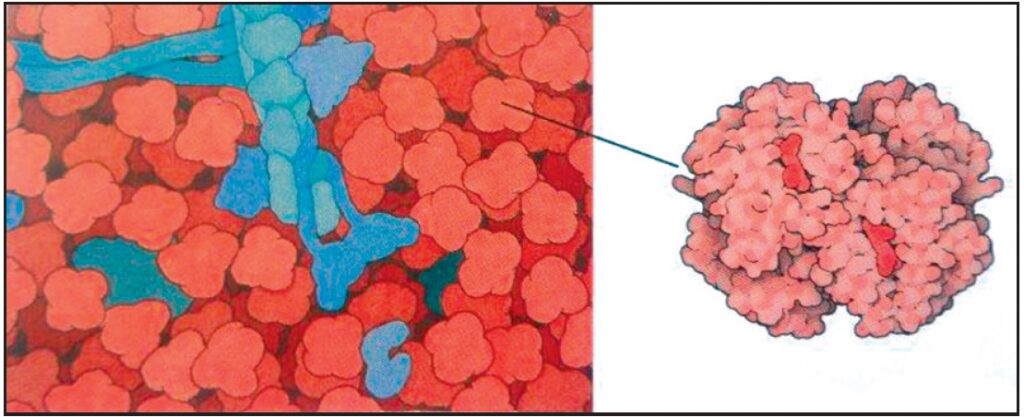
Il nostro sangue contiene proteine che riconoscono e distruggono le cellule e i virus che invadono. Questa immagine mostra una sezione trasversale attraverso una cellula batterica (metà inferiore, in verde, blu e viola) mentre è attaccata da proteine nel siero del sangue (in alto, in giallo e arancione). Anticorpi a forma di Y iniziano il processo legandosi alla superficie della cellula e sono, a loro, volta riconosciuti dalla proteina di sei braccia nella parte centrale superiore. Questo avvia una cascata di azioni che alla fine portano alla formazione di un complesso di attacco alla membrana, qui mostrati mentre penetrano la parete cellulare del batterio (1.000.000 X). D. S. Goodsell. The Machinery of Life, 2009.
I meccanismi molecolari devono anche essere molto specifici per i compiti che svolgono. All’interno di una cellula, un enzima tipico, ad esempio, sarà conteso da migliaia di differenti tipi di molecole. L’enzima deve però essere in grado di cernere attraverso queste numerose distrazioni e afferrare la molecola giusta per guidare (catalizzare) un processo bio-chimico funzionale. Queste proteine enzimatiche svolgono questo compito arduo perché nel tempo si sono evolute per il riconoscimento molecolare. Hanno, inoltre, interazioni molto intime con i loro partner molecolari, entrando in contatto in molti punti e richiedendo una perfetta corrispondenza di forma e chimica. Gli enzimi spesso circondano completamente le loro molecole bersaglio e le proteine si associano attraverso grandi interfacce complementari. A causa di questa notevole specificità, migliaia di reazioni possono accadere allo stesso tempo in una cellula, e il tutto può ancora essere orchestrato per associazione casuale nella brulicante zuppa citoplasmatica41.
In definitiva, cosa dedurre da questi brevi cenni di biologia cellulare?
Che siamo non solo quello che ci appare visibilmente ma, soprattutto, anche quello che non vediamo in un contesto quotidiano, ma scopriamo quando la nostra fisicità da segnali allarmanti: tutto un mondo sottostante che normalmente ignoriamo.
Immagine di copertina magnific.com
- Alberts B., Johnson A., Lewis J., Raff M., Robert K. and Walter P. Molecular Biology of the Cell. Garland Science, New York, 2002.
- Il nanometro (simbolo nm) è un’unità di misura di lunghezza, corrispondente a 10-9 metri (cioè un miliardesimo di metro, pari ad un milionesimo di millimetro)
- Goodsell David.S. Bionanotechnology. Wiley-Liss, Hoboken, 2004.
- Ricordiamo che 1 micron (µm) è la millesima parte di un millimetro (1 micron= 1/1000 di millimetro.
- Goodsell David. S. The Machinery of Life. Springer, New York, 2010, p. 3.
- Le scienze.it
- Alberts B., Johnson A., Lewis J., Raff M., Robert K. and Walter P. op. cit.
- Ora, immaginiamoci l’interazione tra le macromolecole in un ambiente intracellulare. L’interno di una cellula è un ambiente molto complesso e affollato dove le interazioni avvengono caoticamente ma, secondo schemi studiati dalle simulazioni dei ricercatori, certamente, il comportamento molecolare intracellulare è molto diverso da quello studiato in vitro. Ad esempio, all’interno del citoplasma batterico si arriva a notevoli concentrazioni (fino a 400 mg/ml), che modificano sostanzialmente il comportamento di tutte le macromolecole, rispetto a quello delle stesse sostanze quando vengono studiate “in vitro”. Infatti, mentre in provetta le macromolecole in soluzione sono libere di muoversi e di reagire le une con le altre nel solvente, il volume occupato da ciascuna all’interno della membrana citoplasmatica riduce sensibilmente lo spazio e la libertà di movimento di cui godono le altre.
- Alberts B., Johnson A., Lewis J., Raff M., Robert K. and Walter P. op. cit.
- Goodsell David. S. op. cit. 2010, pag. 6.
- La ionizzazione, ad esempio consiste nella generazione di uno o più ioni a causa della rimozione o addizione di elettroni da una entità molecolare neutra (cioè atomi o molecole) che può essere causata da collisioni tra particelle o per assorbimento di radiazioni. Gli atomi o le molecole che hanno un numero di elettroni minore del numero atomico, rimangono carichi positivamente e prendono il nome di “cationi” mentre quelli che hanno un numero di elettroni maggiore del numero atomico, rimangono carichi negativamente e prendono il nome di “anioni”.
- Goodsell David.S. Bionanotechnology. Wiley-Liss, Hoboken, 2004.
- La ATP sintasi (o ATPasi trasportante H+ tra due settori o ATP sintasi traslocante i protoni, o ATP sintasi FoF1 o FoF1 ATPasi o H+-ATP sintasi di tipo F o Complesso V) è un complesso enzimatico che catalizza la seguente reazione: ATP + H2O + H+ interno = ADP + fosfato + H+ esterno Quando la reazione è catalizzata verso sinistra, l’enzima è comunemente chiamato ATP-sintasi ed è responsabile della sintesi di adenosintrifosfato (ATP) utilizzando come substrati adenosindifosfato (ADP) e fosfato inorganico, sfruttando il gradiente protonico generato dalla catena di trasporto degli elettroni. Negli eucarioti, l’enzima è costituito da almeno 16 subunità diverse, alcune in molteplici copie; nei procarioti il numero di subunità è inferiore.
- Goodsell David. S. op. cit. 2010, p. 9-10.
- Pollard Thomas.D. and Earnshaw William .C. Cell Biology. Saunders, Philadelphia, 2002.
- Complementarità chimica intermolecolare – Con il termine generico di legami intermolecolari si intendono le forze attrattive delle molecole. Tali interazioni riguardano sia molecole polari che non polari e sono conosciute, genericamente, come forze di Van der Waals. Le forze di Van der Waals sono forze molto deboli e hanno una energia mediamente cento volte inferiore a quelle di un legame covalente o ionico ( forze intramolecolari), e la loro intensità diminuisce rapidamente con la distanza.
- Alberts B., Johnson A., Lewis J., Raff M., Robert K. and Walter P. Molecular Biology of the Cell. Garland Science, New York, 2002.
- Wikipedia. Legame a idrogeno.
- Ibidem
- Ibidem
- Pollard Thomas.D. and Earnshaw William .C. Cell Biology. Saunders, Philadelphia, 2002.
- Ibidem
- In genetica, una delle due catene polinucleotidiche di una molecola di DNA, che normalmente è costituita da una coppia di filamenti tra loro complementari.
- Ibidem
- Ibidem
- Ibidem
- Alberts B., Johnson A., Lewis J., Raff M., Robert K. and Walter P. Molecular Biology of the Cell. Garland Science, New York, 2002.
- Ibidem & Pollard Thomas.D. and Earnshaw William .C. Cell Biology. Saunders, Philadelphia, 2002.
- Ibidem
- Ibidem
- Ibidem
- Ibidem
- Alberts B., Johnson A., Lewis J., Raff M., Robert K. and Walter P. Molecular Biology of the Cell. Garland Science, New York, 2002.
- Ibidem
- Ibidem
- Ibidem
- Ibidem
- Goodsell David. S. The Machinery of Life. Springer, New York, 2010, p. 25.
- Alberts B., Johnson A., Lewis J., Raff M., Robert K. and Walter P. Molecular Biology of the Cell. Garland Science, New York, 2002 & Pollard Thomas.D. and Earnshaw William .C. Cell Biology. Saunders, Philadelphia, 2002.
- Ibidem
- Ibidem




